

Targeted eradication of EBV-positive cancer cells by CRISPR/dCas9-mediated EBV reactivation in combination with ganciclovir
- PMID: 38874417
- PMCID: PMC11253602
- DOI: 10.1128/mbio.00795-24
Targeted eradication of EBV-positive cancer cells by CRISPR/dCas9-mediated EBV reactivation in combination with ganciclovir
Abstract
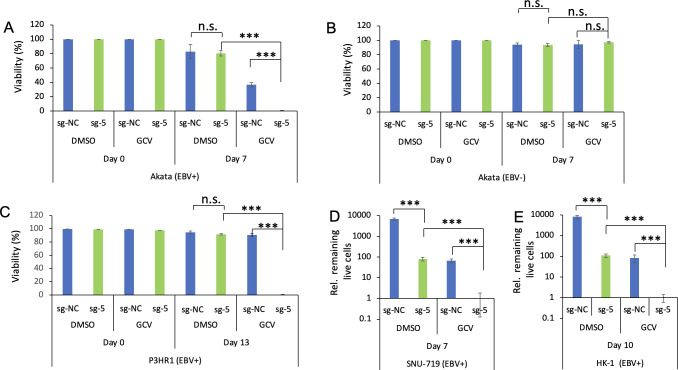
Epstein-Barr virus (EBV) is a ubiquitous human tumor virus that establishes lifelong, persistent infections in B cells. The presence of EBV in cancer cells presents an opportunity to target these cells by reactivating the virus from latency. In this study, we developed a novel approach for EBV reactivation termed clustered regularly interspaced short palindromic repeats (CRISPR)/dCas9-mediated EBV reactivation (CMER) strategy. Using modified CRISPR-associated protein 9 (dCas9) fused with VP64, we designed 10 single guide RNAs (sgRNAs) to target and activate the EBV immediate-early gene promoter. In Akata Burkitt lymphoma cells, 9 out of 10 CMER sgRNAs effectively reactivated EBV. Among these, CMER sgRNA-5 triggered robust reactivation across various cell types, including lymphoma, gastric cancer, and nasopharyngeal carcinoma cells. Importantly, the combination of CMER and ganciclovir selectively eliminated EBV-positive cells, regardless of their cell origin. These findings indicate that targeted virus reactivation by CMER, combined with nucleoside analog therapy, holds promise for EBV-associated cancer treatment.
Importance: This study explores a novel strategy called clustered regularly interspaced short palindromic repeats (CRISPR)/dCas9-mediated Epstein-Barr virus (EBV) reactivation (CMER) to reactivate the Epstein-Barr virus in cancer cells. EBV is associated with various cancers, and reactivating EBV from latency offers a potential therapeutic strategy. We utilized an enzymatically inactive CRISPR-associated protein 9 (dCas9) fused with VP64 and designed 10 single guide RNAs to target the EBV immediate-early gene promoter. Nine of these sgRNAs effectively reactivated EBV in Burkitt lymphoma cells, with CMER sgRNA-5 demonstrating strong reactivation across different cancer cell types. Combining CMER with ganciclovir selectively eliminated EBV-positive cells, showing promise for EBV-associated cancer treatment.
Keywords: CMER; CRISPR/dCas9 activation; EBV; ganciclovir; gastric cancer; latency; lymphoma; nasopharyngeal carcinoma; reactivation; targeted therapy.
Conflict of interest statement
The application for a patent related to this study has been filed by the University of Pittsburgh and Virginia Commonwealth University. R.L. and F.G.S. are inventors on this provisional patent application.
Figures









Similar articles
-
Interferon-γ-inducible protein 16 (IFI16) is required for the maintenance of Epstein-Barr virus latency.Virol J. 2017 Nov 13;14(1):221. doi: 10.1186/s12985-017-0891-5. Virol J. 2017. PMID: 29132393 Free PMC article.
-
Potent and Targeted Activation of Latent HIV-1 Using the CRISPR/dCas9 Activator Complex.Mol Ther. 2016 Mar;24(3):488-98. doi: 10.1038/mt.2015.202. Epub 2015 Nov 19. Mol Ther. 2016. PMID: 26581162 Free PMC article.
-
MicroRNA-focused CRISPR/Cas9 screen identifies miR-142 as a key regulator of Epstein-Barr virus reactivation.PLoS Pathog. 2024 Jun 17;20(6):e1011970. doi: 10.1371/journal.ppat.1011970. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38885264 Free PMC article.
-
Epstein--Barr virus post-transplant lymphoproliferative disease and virus-specific therapy: pharmacological re-activation of viral target genes with arginine butyrate.Transpl Infect Dis. 2001 Sep;3(3):177-85. doi: 10.1034/j.1399-3062.2001.003003177.x. Transpl Infect Dis. 2001. PMID: 11493400 Review.
-
Reactivation of Epstein-Barr virus from latency.Rev Med Virol. 2005 May-Jun;15(3):149-56. doi: 10.1002/rmv.456. Rev Med Virol. 2005. PMID: 15546128 Review.
Cited by
-
Leveraging the interconnected unfolded protein response and NLRP3 inflammasome pathways to reactivate Epstein-Barr virus in diffuse large B-cell lymphomas.NAR Cancer. 2025 May 5;7(2):zcaf017. doi: 10.1093/narcan/zcaf017. eCollection 2025 Jun. NAR Cancer. 2025. PMID: 40330549 Free PMC article.
-
Targeting EBV Episome for Anti-Cancer Therapy: Emerging Strategies and Challenges.Viruses. 2025 Jan 15;17(1):110. doi: 10.3390/v17010110. Viruses. 2025. PMID: 39861899 Free PMC article. Review.
-
Applying CRISPR Technologies for the Treatment of Human Herpesvirus Infections: A Scoping Review.Pathogens. 2025 Jul 1;14(7):654. doi: 10.3390/pathogens14070654. Pathogens. 2025. PMID: 40732701 Free PMC article. Review.
-
CRISPR/Cas13-Mediated Inhibition of EBNA1 for Suppression of Epstein-Barr Virus Transcripts and DNA Load in Nasopharyngeal Carcinoma Cells.Viruses. 2025 Jun 26;17(7):899. doi: 10.3390/v17070899. Viruses. 2025. PMID: 40733516 Free PMC article.
References
-
- Messick TE, Smith GR, Soldan SS, McDonnell ME, Deakyne JS, Malecka KA, Tolvinski L, van den Heuvel APJ, Gu B-W, Cassel JA, Tran DH, Wassermann BR, Zhang Y, Velvadapu V, Zartler ER, Busson P, Reitz AB, Lieberman PM. 2019. Structure-based design of small-molecule inhibitors of EBNA1 DNA binding blocks Epstein-Barr virus latent infection and tumor growth. Sci Transl Med 11:eaau5612. doi:10.1126/scitranslmed.aau5612 - DOI - PMC - PubMed
-
- Meng Q, Hagemeier SR, Fingeroth JD, Gershburg E, Pagano JS, Kenney SC. 2010. The Epstein-Barr virus (EBV)-encoded protein kinase, EBV-PK, but not the thymidine kinase (EBV-TK), is required for ganciclovir and acyclovir inhibition of lytic viral production. J Virol 84:4534–4542. doi:10.1128/JVI.02487-09 - DOI - PMC - PubMed
-
- Westphal EM, Mauser A, Swenson J, Davis MG, Talarico CL, Kenney SC. 1999. Induction of lytic Epstein-Barr virus (EBV) infection in EBV-associated malignancies using adenovirus vectors in vitro and in vivo. Cancer Res 59:1485–1491. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
