

Cooperative Gsx2-DNA binding requires DNA bending and a novel Gsx2 homeodomain interface
- PMID: 38874471
- PMCID: PMC11260452
- DOI: 10.1093/nar/gkae522
Cooperative Gsx2-DNA binding requires DNA bending and a novel Gsx2 homeodomain interface
Erratum in
-
Correction to 'Cooperative Gsx2-DNA binding requires DNA bending and a novel Gsx2 homeodomain interface'.Nucleic Acids Res. 2024 Jul 22;52(13):8037. doi: 10.1093/nar/gkae578. Nucleic Acids Res. 2024. PMID: 38932680 Free PMC article. No abstract available.
Abstract
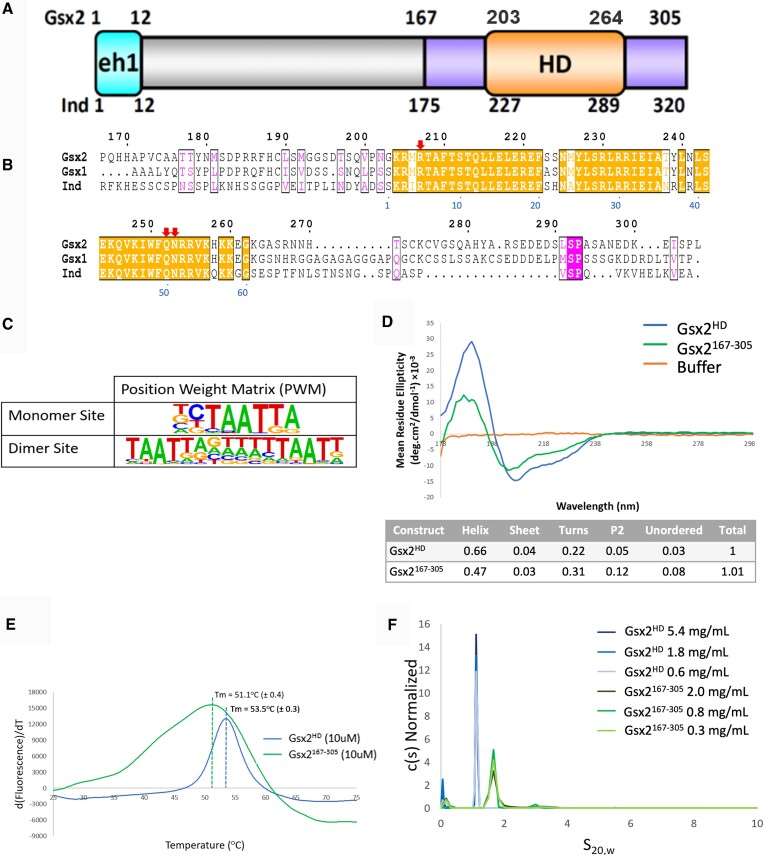
The conserved Gsx homeodomain (HD) transcription factors specify neural cell fates in animals from flies to mammals. Like many HD proteins, Gsx factors bind A/T-rich DNA sequences prompting the following question: How do HD factors that bind similar DNA sequences in vitro regulate specific target genes in vivo? Prior studies revealed that Gsx factors bind DNA both as a monomer on individual A/T-rich sites and as a cooperative homodimer to two sites spaced precisely 7 bp apart. However, the mechanistic basis for Gsx-DNA binding and cooperativity is poorly understood. Here, we used biochemical, biophysical, structural and modeling approaches to (i) show that Gsx factors are monomers in solution and require DNA for cooperative complex formation, (ii) define the affinity and thermodynamic binding parameters of Gsx2/DNA interactions, (iii) solve a high-resolution monomer/DNA structure that reveals that Gsx2 induces a 20° bend in DNA, (iv) identify a Gsx2 protein-protein interface required for cooperative DNA binding and (v) determine that flexible spacer DNA sequences enhance Gsx2 cooperativity on dimer sites. Altogether, our results provide a mechanistic basis for understanding the protein and DNA structural determinants that underlie cooperative DNA binding by Gsx factors.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






Update of
-
Cooperative Gsx2-DNA Binding Requires DNA Bending and a Novel Gsx2 Homeodomain Interface.bioRxiv [Preprint]. 2023 Dec 8:2023.12.08.570805. doi: 10.1101/2023.12.08.570805. bioRxiv. 2023. Update in: Nucleic Acids Res. 2024 Jul 22;52(13):7987-8002. doi: 10.1093/nar/gkae522. PMID: 38106145 Free PMC article. Updated. Preprint.
References
-
- Pick L., Heffer A. Hox gene evolution: multiple mechanisms contributing to evolutionary novelties. Ann. N.Y. Acad. Sci. 2012; 1256:15–32. - PubMed
-
- Jolma A., Yan J., Whitington T., Toivonen J., Nitta K.R., Rastas P., Morgunova E., Enge M., Taipale M., Wei G. et al. DNA-binding specificities of human transcription factors. Cell. 2013; 152:327–339. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
