

Genome-wide association studies meta-analysis uncovers NOJO and SGS3 novel genes involved in Arabidopsis thaliana primary root development and plasticity
- PMID: 38874813
- PMCID: PMC11178574
- DOI: 10.1007/s11033-024-09623-1
Genome-wide association studies meta-analysis uncovers NOJO and SGS3 novel genes involved in Arabidopsis thaliana primary root development and plasticity
Abstract
Background: Arabidopsis thaliana primary root growth has become a model for evo-devo studies due to its simplicity and facility to record cell proliferation and differentiation. To identify new genetic components relevant to primary root growth, we used a Genome-Wide Association Studies (GWAS) meta-analysis approach using data published in the last decade. In this work, we performed intra and inter-studies analyses to discover new genetic components that could participate in primary root growth.
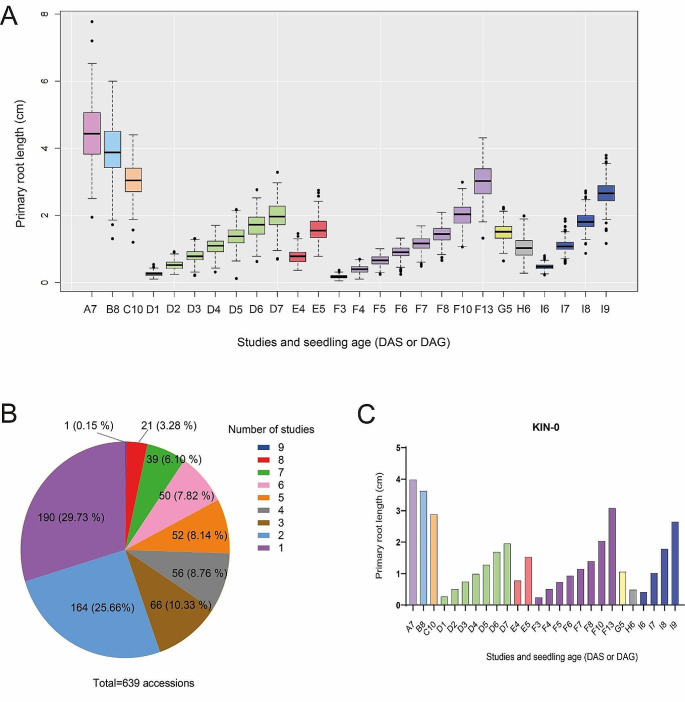
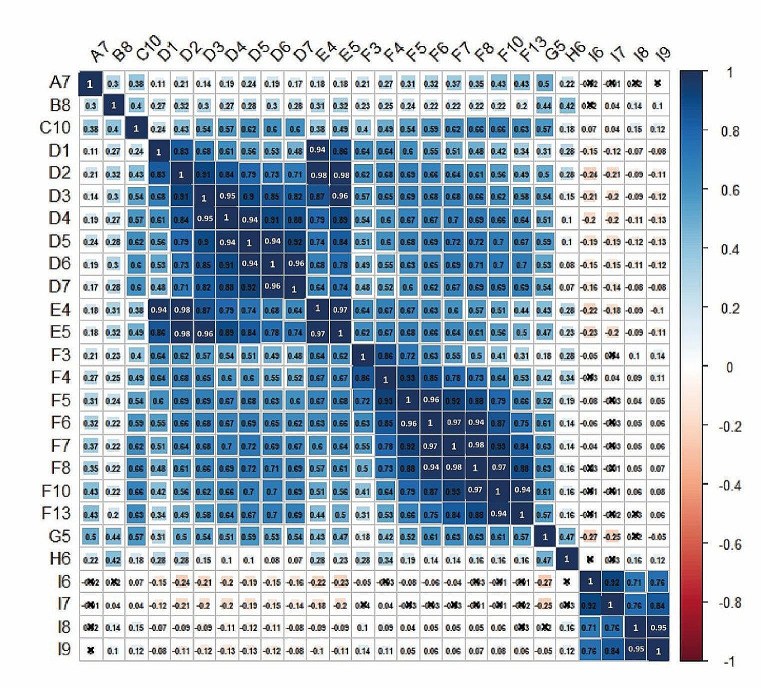
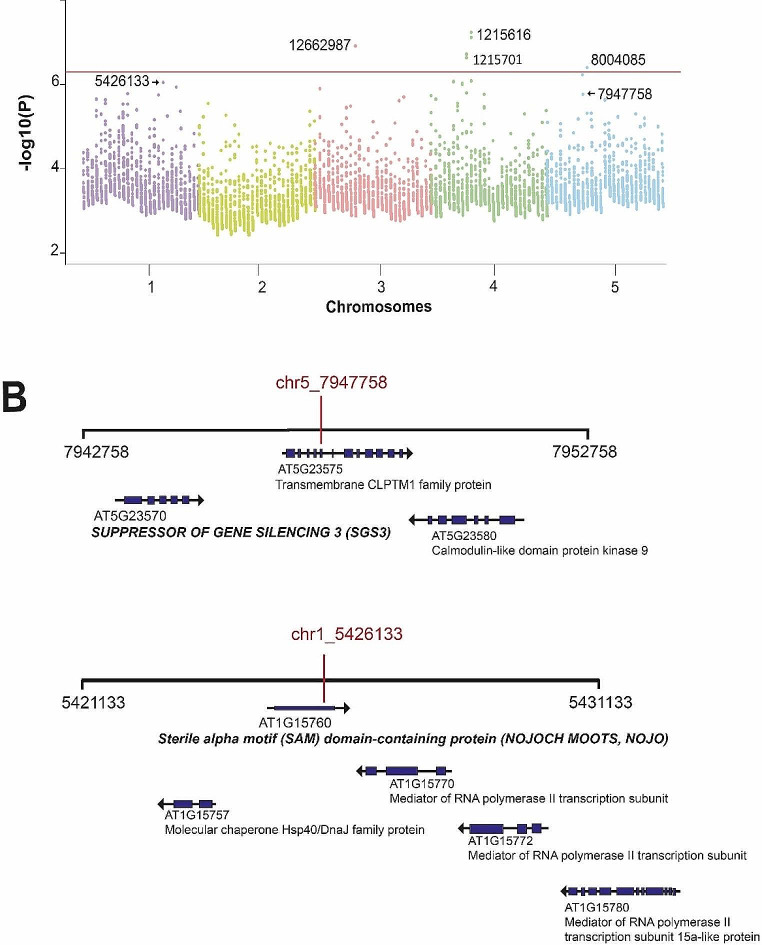
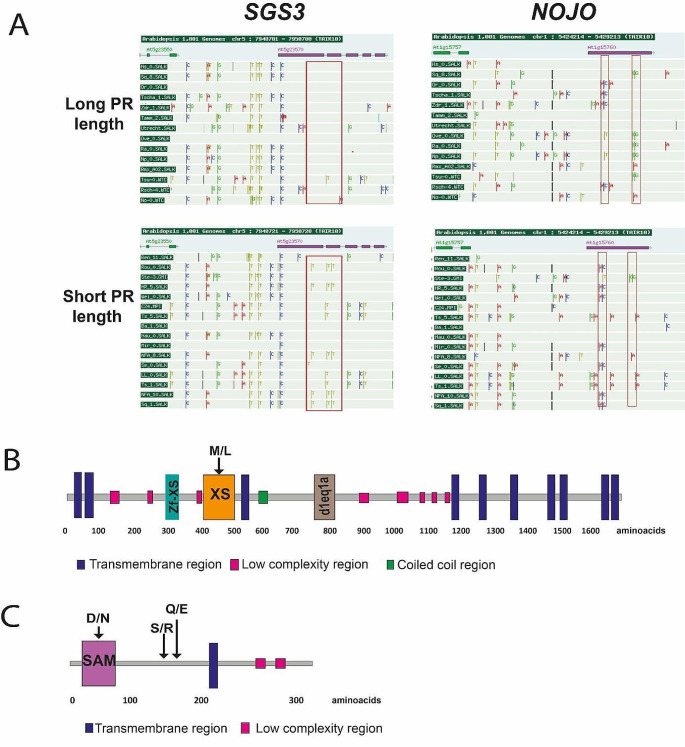
Methods and results: We used 639 accessions from nine different studies under control conditions and performed different GWAS tests. We found that primary root growth changes were associated with 41 genes, of which six (14.6%) have been previously described as inhibitors or promoters of primary root growth. The knockdown lines of two genes, Suppressor of Gene Silencing (SGS3), involved in tasiRNA processing, and a gene with a Sterile Alpha Motif (SAM) motif named NOJOCH MOOTS (NOJO), confirmed their role as repressors of primary root growth, none has been shown to participate in this developmental process before.
Conclusions: In summary, our GWAS analysis of different available studies identified new genes that participate in primary root growth; two of them were identified as repressors of primary root growth.
Keywords: Arabidopsis thaliana; NOJO; SGS3; Accessions; GWAS; Primary root growth.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Meinke DW, Cherry JM, Dean C et al (1998) Arabidopsis thaliana: a Model Plant for Genome Analysis. Science 282:662–682. 10.1126/science.282.5389.662 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
