

Microbiota dictate T cell clonal selection to augment graft-versus-host disease after stem cell transplantation
- PMID: 38876098
- PMCID: PMC11236519
- DOI: 10.1016/j.immuni.2024.05.018
Microbiota dictate T cell clonal selection to augment graft-versus-host disease after stem cell transplantation
Abstract
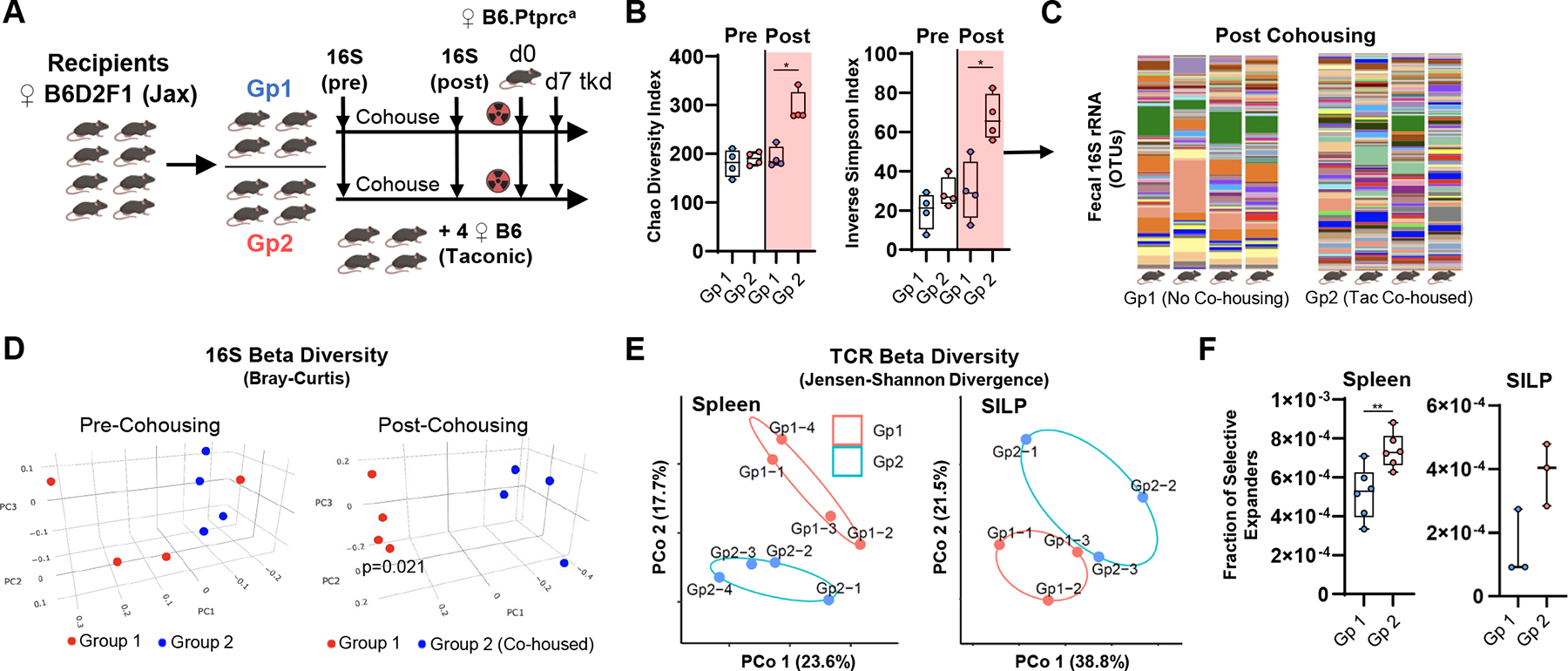
Allogeneic T cell expansion is the primary determinant of graft-versus-host disease (GVHD), and current dogma dictates that this is driven by histocompatibility antigen disparities between donor and recipient. This paradigm represents a closed genetic system within which donor T cells interact with peptide-major histocompatibility complexes (MHCs), though clonal interrogation remains challenging due to the sparseness of the T cell repertoire. We developed a Bayesian model using donor and recipient T cell receptor (TCR) frequencies in murine stem cell transplant systems to define limited common expansion of T cell clones across genetically identical donor-recipient pairs. A subset of donor CD4+ T cell clonotypes differentially expanded in identical recipients and were microbiota dependent. Microbiota-specific T cells augmented GVHD lethality and could target microbial antigens presented by gastrointestinal epithelium during an alloreactive response. The microbiota serves as a source of cognate antigens that contribute to clonotypic T cell expansion and the induction of GVHD independent of donor-recipient genetics.
Keywords: CD4(+) T cell; GVHD; T cell repertoire; graft-versus-host disease; histocompatibility; microbiome; microbiota T cells; stem cell transplant.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.R.H. has consulted for Generon Corporation, NapaJen Pharma, iTeos Therapeutics, and Neoleukin Therapeutics and has received unrelated research funding from Compass Therapeutics, Syndax Pharmaceuticals, Applied Molecular Transport, Serplus Technology, Heat Biologics, Laevoroc Oncology, iTeos Therapeutics, Genentech, and CSL Behring.
Figures






References
-
- Goulmy E, Schipper R, Pool J, Blokland E, Falkenburg JH, Vossen J, Gratwohl A, Vogelsang GB, van Houwelingen HC, and van Rood JJ (1996). Mismatches of minor histocompatibility antigens between HLA-identical donors and recipients and the development of graft-versus-host disease after bone marrow transplantation. N. Engl. J. Med 334, 281–285. 10.1056/NEJM199602013340501. - DOI - PubMed
-
- Meier J, Roberts C, Avent K, Hazlett A, Berrie J, Payne K, Hamm D, Desmarais C, Sanders C, Hogan KT, et al. (2013). Fractal organization of the human T cell repertoire in health and after stem cell transplantation. Biol. Blood Marrow Transplant 19, 366–377. 10.1016/j.bbmt.2012.12.004. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
