

Uncovering the dynamics of precise repair at CRISPR/Cas9-induced double-strand breaks
- PMID: 38877047
- PMCID: PMC11178868
- DOI: 10.1038/s41467-024-49410-x
Uncovering the dynamics of precise repair at CRISPR/Cas9-induced double-strand breaks
Abstract
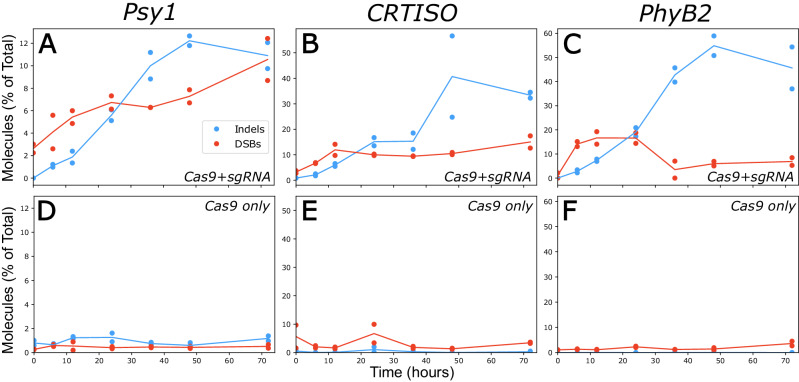
CRISPR/Cas9 is widely used for precise mutagenesis through targeted DNA double-strand breaks (DSBs) induction followed by error-prone repair. A better understanding of this process requires measuring the rates of cutting, error-prone, and precise repair, which have remained elusive so far. Here, we present a molecular and computational toolkit for multiplexed quantification of DSB intermediates and repair products by single-molecule sequencing. Using this approach, we characterize the dynamics of DSB induction, processing and repair at endogenous loci along a 72 h time-course in tomato protoplasts. Combining this data with kinetic modeling reveals that indel accumulation is determined by the combined effect of the rates of DSB induction processing of broken ends, and precise versus error repair. In this study, 64-88% of the molecules were cleaved in the three targets analyzed, while indels ranged between 15-41%. Precise repair accounts for most of the gap between cleavage and error repair, representing up to 70% of all repair events. Altogether, this system exposes flux in the DSB repair process, decoupling induction and repair dynamics, and suggesting an essential role of high-fidelity repair in limiting the efficiency of CRISPR-mediated mutagenesis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







Similar articles
-
Kinetics and Fidelity of the Repair of Cas9-Induced Double-Strand DNA Breaks.Mol Cell. 2018 Jun 7;70(5):801-813.e6. doi: 10.1016/j.molcel.2018.04.016. Epub 2018 May 24. Mol Cell. 2018. PMID: 29804829 Free PMC article.
-
INDEL detection, the 'Achilles heel' of precise genome editing: a survey of methods for accurate profiling of gene editing induced indels.Nucleic Acids Res. 2020 Dec 2;48(21):11958-11981. doi: 10.1093/nar/gkaa975. Nucleic Acids Res. 2020. PMID: 33170255 Free PMC article. Review.
-
Single-Strand Annealing Plays a Major Role in Double-Strand DNA Break Repair following CRISPR-Cas9 Cleavage in Leishmania.mSphere. 2019 Aug 21;4(4):e00408-19. doi: 10.1128/mSphere.00408-19. mSphere. 2019. PMID: 31434745 Free PMC article.
-
Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions.Genetics. 2013 Oct;195(2):331-48. doi: 10.1534/genetics.113.155382. Epub 2013 Aug 9. Genetics. 2013. PMID: 23934893 Free PMC article.
-
Mapping cellular responses to DNA double-strand breaks using CRISPR technologies.Trends Genet. 2023 Jul;39(7):560-574. doi: 10.1016/j.tig.2023.02.015. Epub 2023 Mar 25. Trends Genet. 2023. PMID: 36967246 Free PMC article. Review.
Cited by
-
Dual activities of an X-family DNA polymerase regulate CRISPR-induced insertional mutagenesis across species.Nat Commun. 2024 Jul 26;15(1):6293. doi: 10.1038/s41467-024-50676-4. Nat Commun. 2024. PMID: 39060288 Free PMC article.
-
Widespread position-dependent transcriptional regulatory sequences in plants.Nat Genet. 2024 Oct;56(10):2238-2246. doi: 10.1038/s41588-024-01907-3. Epub 2024 Sep 12. Nat Genet. 2024. PMID: 39266765 Free PMC article.
-
Characterization of radiations-induced genomic structural variations in Arabidopsis thaliana.Plant J. 2025 Jan;121(1):e17180. doi: 10.1111/tpj.17180. Epub 2024 Dec 1. Plant J. 2025. PMID: 39616610 Free PMC article.
-
RAD52 and ERCC6L/PICH have a compensatory relationship for genome stability in mitosis.PLoS Genet. 2024 Nov 19;20(11):e1011479. doi: 10.1371/journal.pgen.1011479. eCollection 2024 Nov. PLoS Genet. 2024. PMID: 39561207 Free PMC article.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
