

APP fragment controls both ionotropic and non-ionotropic signaling of NMDA receptors
- PMID: 38878768
- PMCID: PMC11343662
- DOI: 10.1016/j.neuron.2024.05.027
APP fragment controls both ionotropic and non-ionotropic signaling of NMDA receptors
Abstract
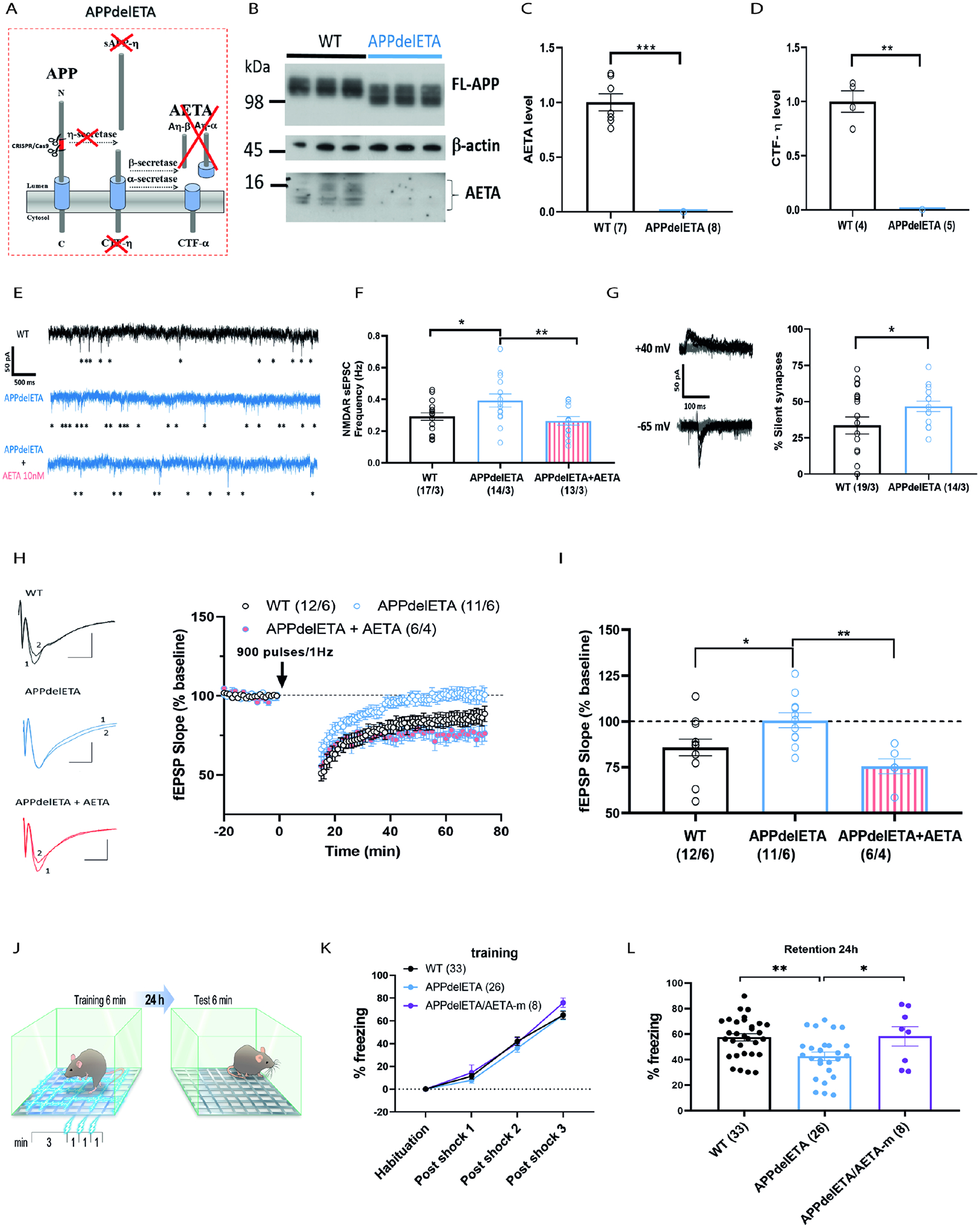
NMDA receptors (NMDARs) are ionotropic receptors crucial for brain information processing. Yet, evidence also supports an ion-flux-independent signaling mode mediating synaptic long-term depression (LTD) and spine shrinkage. Here, we identify AETA (Aη), an amyloid-β precursor protein (APP) cleavage product, as an NMDAR modulator with the unique dual regulatory capacity to impact both signaling modes. AETA inhibits ionotropic NMDAR activity by competing with the co-agonist and induces an intracellular conformational modification of GluN1 subunits. This favors non-ionotropic NMDAR signaling leading to enhanced LTD and favors spine shrinkage. Endogenously, AETA production is increased by in vivo chemogenetically induced neuronal activity. Genetic deletion of AETA production alters NMDAR transmission and prevents LTD, phenotypes rescued by acute exogenous AETA application. This genetic deletion also impairs contextual fear memory. Our findings demonstrate AETA-dependent NMDAR activation (ADNA), characterizing AETA as a unique type of endogenous NMDAR modulator that exerts bidirectional control over NMDAR signaling and associated information processing.
Keywords: APP; NMDA receptors; amyloid-β precursor protein; eta-secretase; hippocampus; long-term depression; memory; non-ionotropic signaling; spine shrinkage; synapse.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
