

The brain microvasculature is a primary mediator of interferon-α neurotoxicity in human cerebral interferonopathies
- PMID: 38878770
- PMCID: PMC11250091
- DOI: 10.1016/j.immuni.2024.05.017
The brain microvasculature is a primary mediator of interferon-α neurotoxicity in human cerebral interferonopathies
Abstract
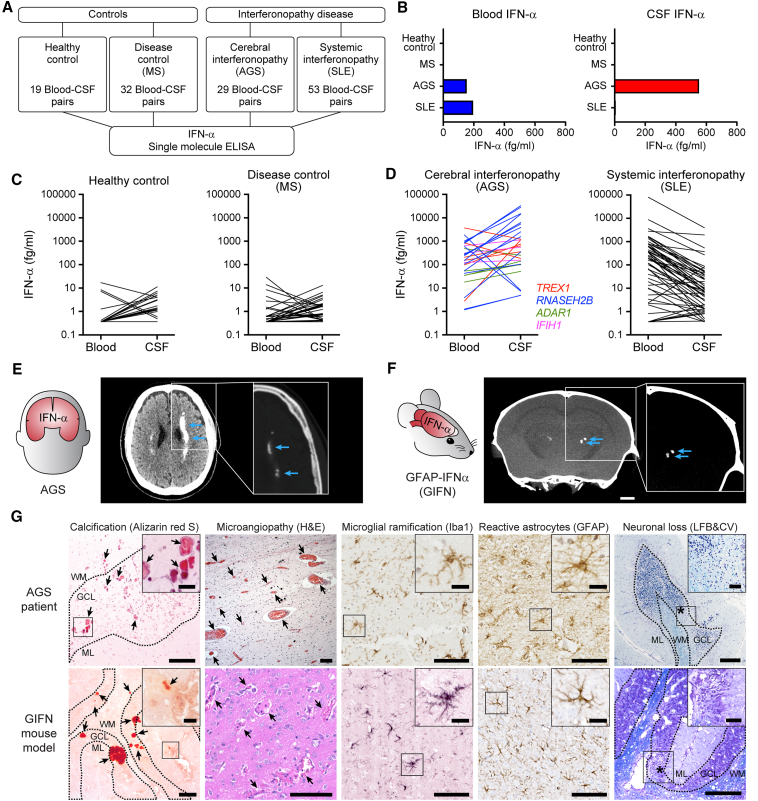
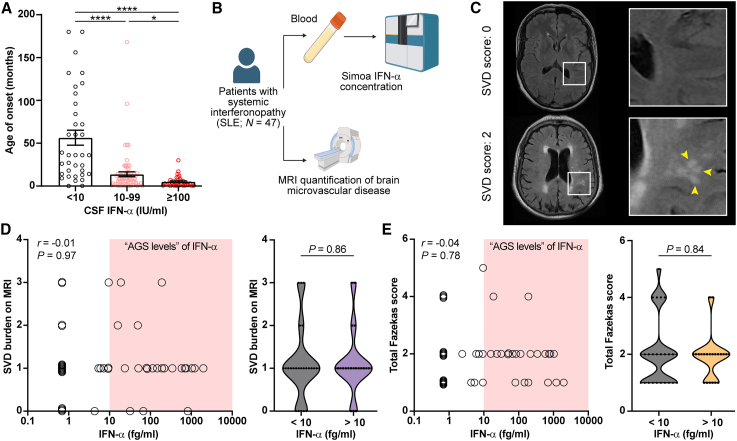
Aicardi-Goutières syndrome (AGS) is an autoinflammatory disease characterized by aberrant interferon (IFN)-α production. The major cause of morbidity in AGS is brain disease, yet the primary source and target of neurotoxic IFN-α remain unclear. Here, we demonstrated that the brain was the primary source of neurotoxic IFN-α in AGS and confirmed the neurotoxicity of intracerebral IFN-α using astrocyte-driven Ifna1 misexpression in mice. Using single-cell RNA sequencing, we demonstrated that intracerebral IFN-α-activated receptor (IFNAR) signaling within cerebral endothelial cells caused a distinctive cerebral small vessel disease similar to that observed in individuals with AGS. Magnetic resonance imaging (MRI) and single-molecule ELISA revealed that central and not peripheral IFN-α was the primary determinant of microvascular disease in humans. Ablation of endothelial Ifnar1 in mice rescued microvascular disease, stopped the development of diffuse brain disease, and prolonged lifespan. These results identify the cerebral microvasculature as a primary mediator of IFN-α neurotoxicity in AGS, representing an accessible target for therapeutic intervention.
Keywords: Aicardi-Goutières syndrome; blood-brain barrier; cerebral interferonopathy; endothelial; interferon-alpha; microangiopathy; neuroinflammation; neurotoxicity; small vessel disease.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests I.L.C. and M.J.H. received funding from Ionis Pharmaceuticals for experiments in an unrelated study using the GIFN mice.
Figures






Comment on
-
Interferon-α receptor antisense oligonucleotides reduce neuroinflammation and neuropathology in a mouse model of cerebral interferonopathy.J Clin Invest. 2024 Jan 9;134(4):e169562. doi: 10.1172/JCI169562. J Clin Invest. 2024. PMID: 38357922 Free PMC article.
References
-
- Crow Y.J., Chase D.S., Lowenstein Schmidt J., Szynkiewicz M., Forte G.M.A., Gornall H.L., Oojageer A., Anderson B., Pizzino A., Helman G., et al. Characterization of human disease phenotypes associated with mutations in TREX1, RNASEH2A, RNASEH2B, RNASEH2C, SAMHD1, ADAR, and IFIH1. Am. J. Med. Genet. A. 2015;167A:296–312. doi: 10.1002/ajmg.a.36887. - DOI - PMC - PubMed
-
- Uggenti C., Lepelley A., Depp M., Badrock A.P., Rodero M.P., El-Daher M.T., Rice G.I., Dhir S., Wheeler A.P., Dhir A., et al. cGAS-mediated induction of type I interferon due to inborn errors of histone pre-mRNA processing. Nat. Genet. 2020;52:1364–1372. doi: 10.1038/s41588-020-00737-3. - DOI - PubMed
-
- Rodero M.P., Decalf J., Bondet V., Hunt D., Rice G.I., Werneke S., McGlasson S.L., Alyanakian M.A., Bader-Meunier B., Barnerias C., et al. Detection of interferon alpha protein reveals differential levels and cellular sources in disease. J. Exp. Med. 2017;214:1547–1555. doi: 10.1084/jem.20161451. - DOI - PMC - PubMed
-
- Rice G.I., Forte G.M.A., Szynkiewicz M., Chase D.S., Aeby A., Abdel-Hamid M.S., Ackroyd S., Allcock R., Bailey K.M., Balottin U., et al. Assessment of interferon-related biomarkers in Aicardi-Goutières syndrome associated with mutations in TREX1, RNASEH2A, RNASEH2B, RNASEH2C, SAMHD1, and ADAR: a case-control study. Lancet Neurol. 2013;12:1159–1169. doi: 10.1016/S1474-4422(13)70258-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
