

Inhibition of WNT/β-catenin signalling during sex-specific gonadal differentiation is essential for normal human fetal testis development
- PMID: 38879537
- PMCID: PMC11180390
- DOI: 10.1186/s12964-024-01704-9
Inhibition of WNT/β-catenin signalling during sex-specific gonadal differentiation is essential for normal human fetal testis development
Abstract
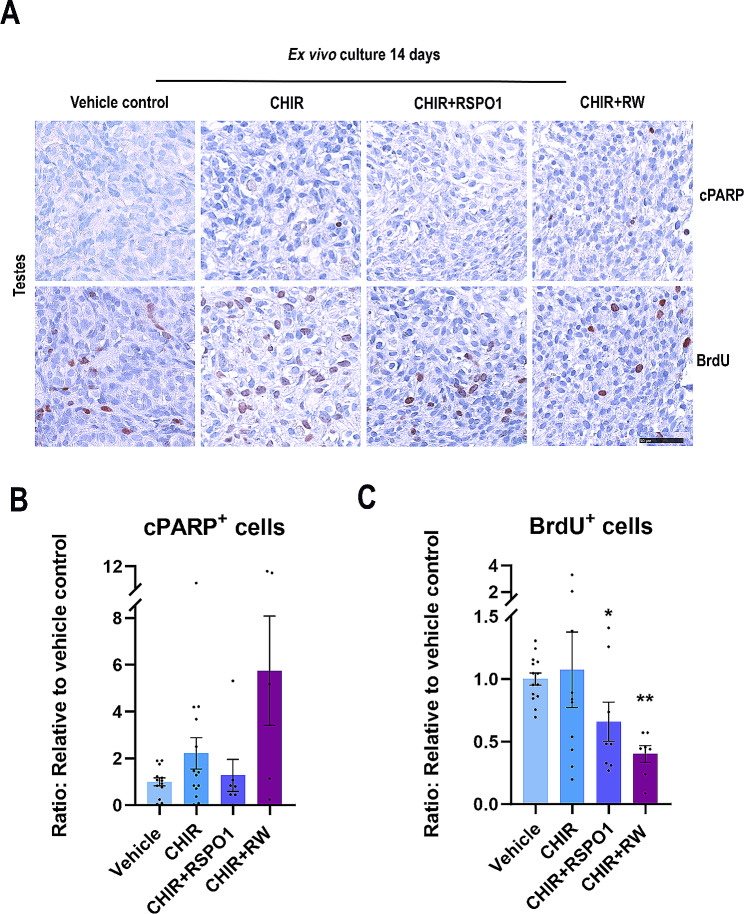
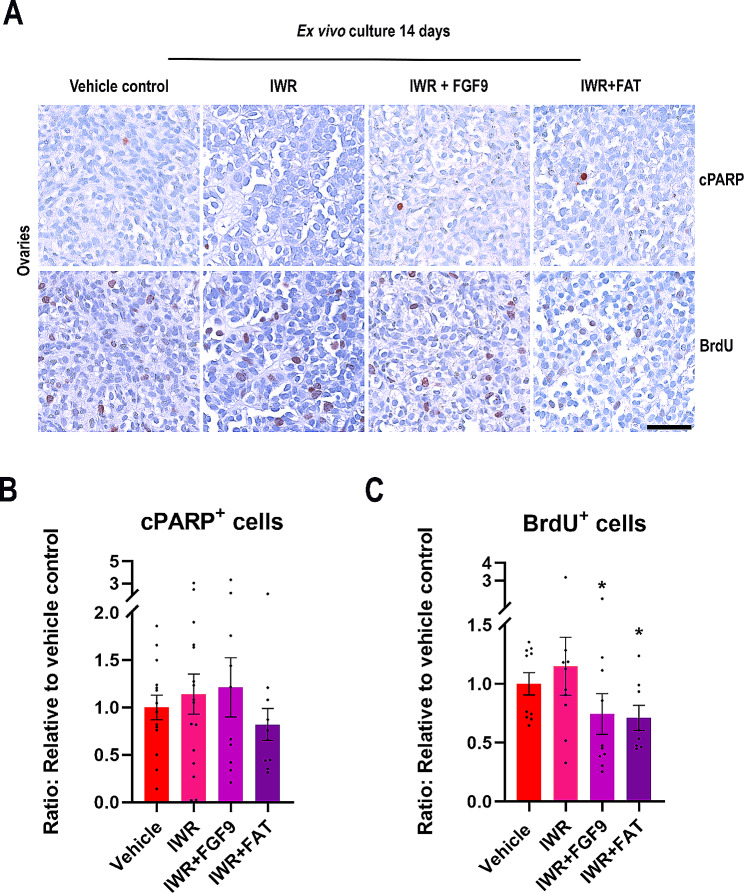
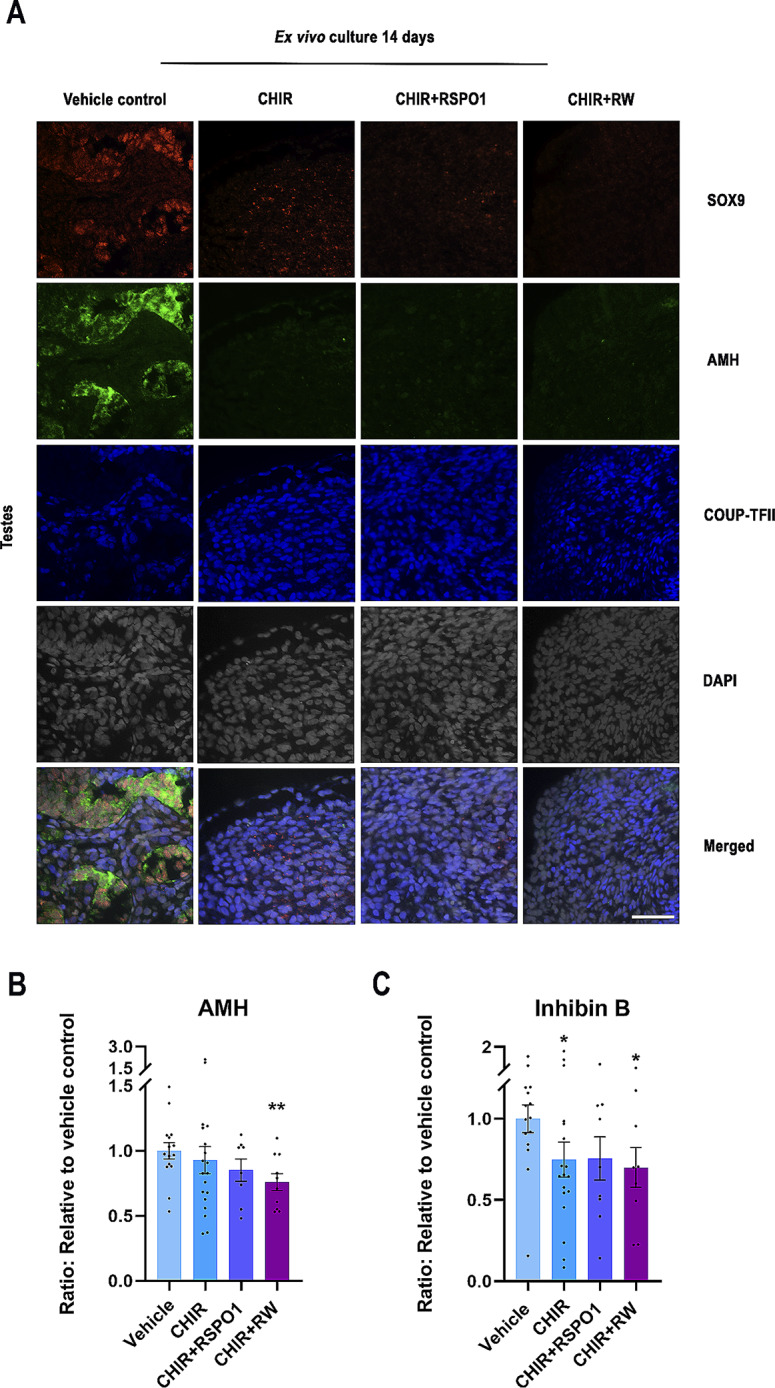
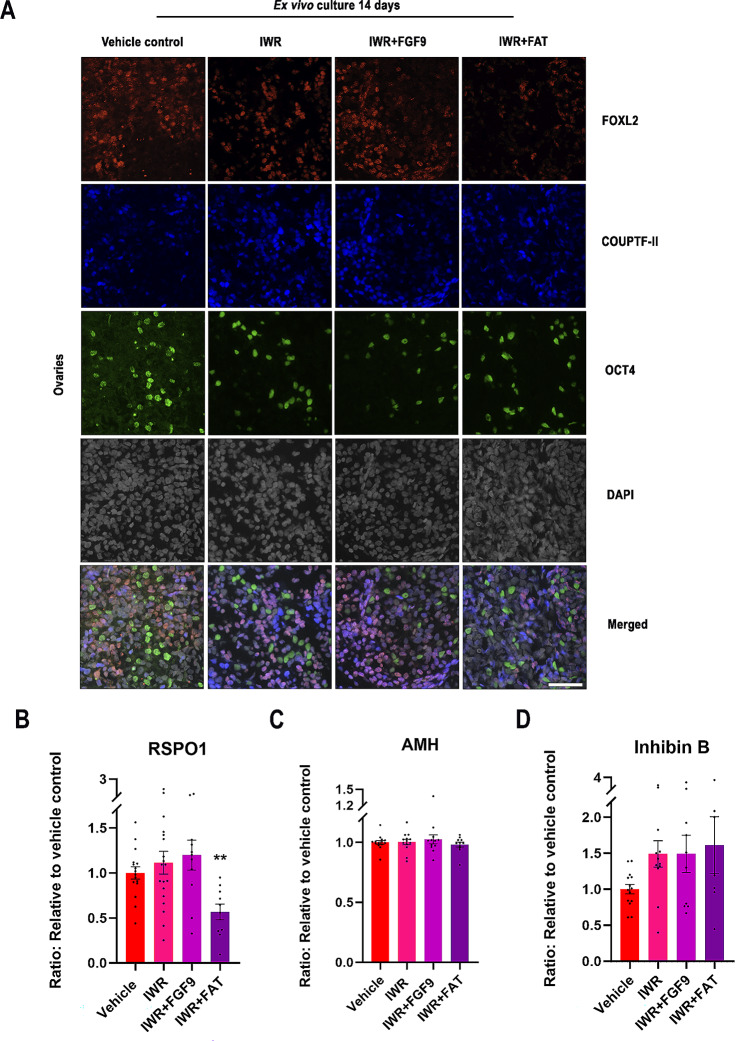
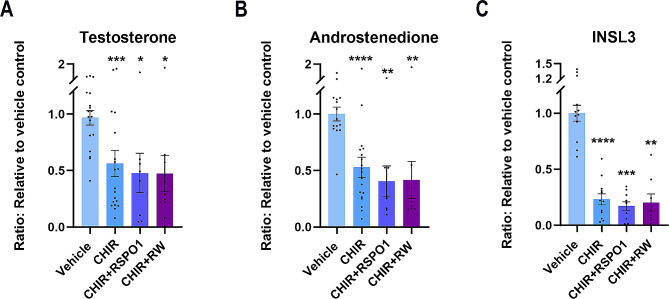
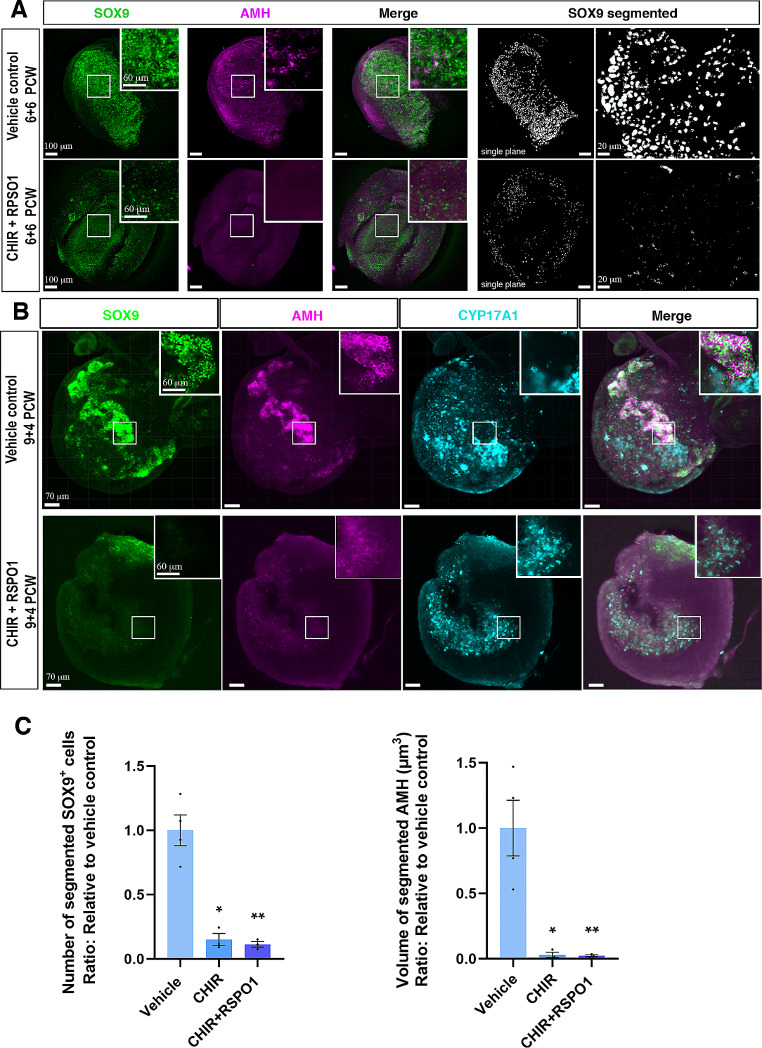
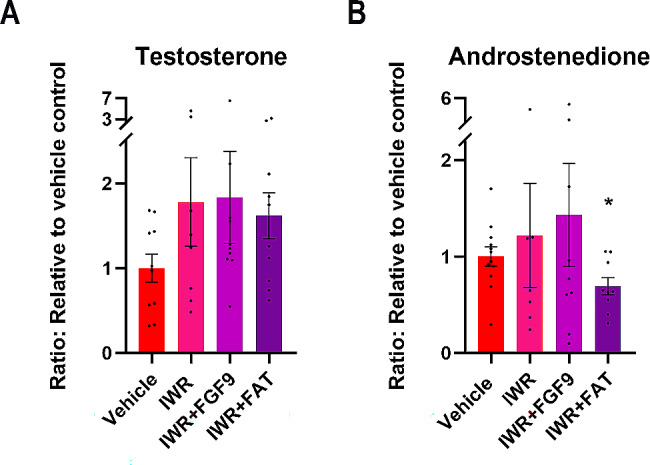
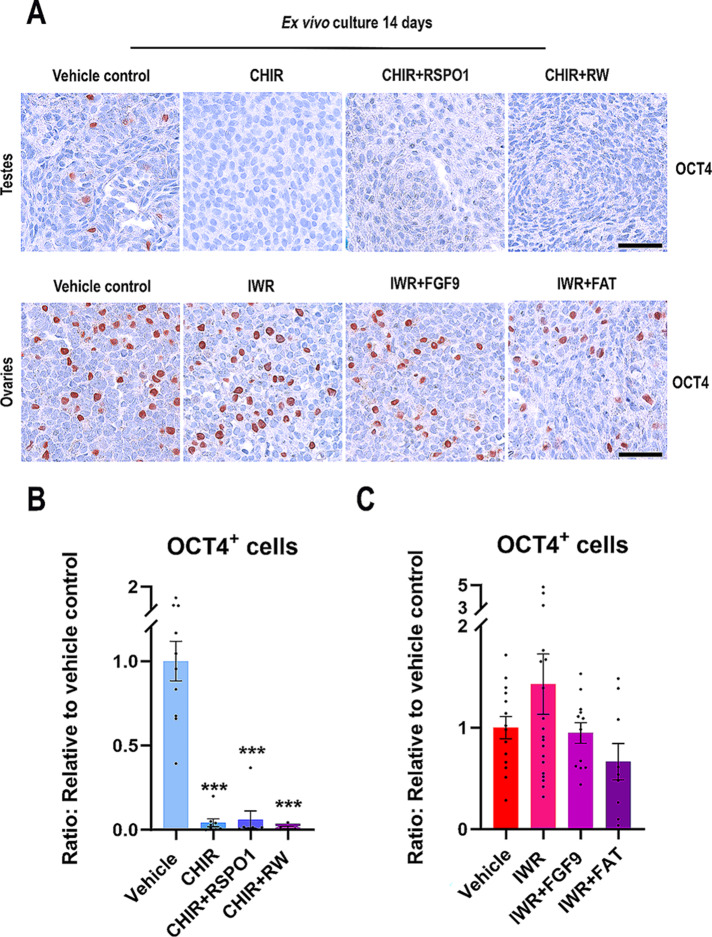
Sex-specific gonadal differentiation is directed by complex signalling promoting development in either male or female direction, while simultaneously inhibiting the opposite pathway. In mice, the WNT/β-catenin pathway promotes ovarian development and the importance of actively inhibiting this pathway to ensure normal testis development has been recognised. However, the implications of alterations in the tightly regulated WNT/β-catenin signalling during human fetal gonad development has not yet been examined in detail. Thus, the aim of this study was to examine the consequences of dysregulating the WNT/β-catenin signalling pathway in the supporting cell lineage during sex-specific human fetal gonad development using an established and extensively validated ex vivo culture model. Inhibition of WNT/β-catenin signalling in human fetal ovary cultures resulted in only minor effects, including reduced secretion of RSPO1 and reduced cell proliferation although this was not consistently found in all treatment groups. In contrast, promotion of WNT/β-catenin signalling in testes severely affected development and function. This included disrupted seminiferous cord structures, reduced cell proliferation, reduced expression of SOX9/AMH, reduced secretion of Inhibin B and AMH as well as loss of the germ cell population. Additionally, Leydig cell function was markedly impaired with reduced secretion of testosterone, androstenedione and INSL3. Together, this study suggests that dysregulated WNT/β-catenin signalling during human fetal gonad development severely impairs testicular development and function. Importantly, our study highlights the notion that sufficient inhibition of the opposite pathway during sex-specific gonadal differentiation is essential to ensure normal development and function also applies to human fetal gonads.
Keywords: Ex vivo culture; Germ cell development; Human fetal gonads; Ovarian and testicular differentiation; Sex-specific development; Supporting cell lineages; WNT/β-catenin signalling.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Dysregulation of FGFR signalling by a selective inhibitor reduces germ cell survival in human fetal gonads of both sexes and alters the somatic niche in fetal testes.Hum Reprod. 2019 Nov 1;34(11):2228-2243. doi: 10.1093/humrep/dez191. Hum Reprod. 2019. PMID: 31734698 Free PMC article.
-
Characterization of somatic testicular cells during human development: fetal, peripubertal, adolescent and adult human testis from healthy and infertility related disease.Int J Dev Biol. 2025 Jun 13;69(2):91-99. doi: 10.1387/ijdb.250005ce. Int J Dev Biol. 2025. PMID: 40521686
-
Transcriptome analysis reveals the sex-switching mechanism of juvenile hermaphroditism in silver pomfret (Pampus argenteus).Biol Sex Differ. 2025 Jul 14;16(1):51. doi: 10.1186/s13293-025-00736-1. Biol Sex Differ. 2025. PMID: 40660379 Free PMC article.
-
Stem cell insights into human trophoblast lineage differentiation.Hum Reprod Update. 2016 Dec;23(1):77-103. doi: 10.1093/humupd/dmw026. Epub 2016 Sep 2. Hum Reprod Update. 2016. PMID: 27591247
-
Impact of residual disease as a prognostic factor for survival in women with advanced epithelial ovarian cancer after primary surgery.Cochrane Database Syst Rev. 2022 Sep 26;9(9):CD015048. doi: 10.1002/14651858.CD015048.pub2. Cochrane Database Syst Rev. 2022. PMID: 36161421 Free PMC article.
Cited by
-
The molecule events expression of TGF-β/Smad signaling pathway in morphological and structural developmental characteristics of gonads in goose embryos.Poult Sci. 2025 Feb;104(2):104808. doi: 10.1016/j.psj.2025.104808. Epub 2025 Jan 11. Poult Sci. 2025. PMID: 39823842 Free PMC article.
-
Identification and profiling of microRNAs during sheep's testicular development.Front Vet Sci. 2025 Mar 31;12:1538990. doi: 10.3389/fvets.2025.1538990. eCollection 2025. Front Vet Sci. 2025. PMID: 40230794 Free PMC article.
-
Genetic and genomic insights into male reproductive tract development.Fertil Steril. 2025 Jun;123(6):970-979. doi: 10.1016/j.fertnstert.2025.03.024. Epub 2025 Mar 31. Fertil Steril. 2025. PMID: 40174856
-
CCN5 Drives Leydig Cell Aging and Testicular Dysfunction: Insights into Fibrosis, Lipid Dysregulation, and Therapeutic Potential.Research (Wash D C). 2025 Aug 1;8:0762. doi: 10.34133/research.0762. eCollection 2025. Research (Wash D C). 2025. PMID: 40756763 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
