

A novel prolixicin identified in common bed bugs with activity against both bacteria and parasites
- PMID: 38879638
- PMCID: PMC11180110
- DOI: 10.1038/s41598-024-64691-4
A novel prolixicin identified in common bed bugs with activity against both bacteria and parasites
Abstract
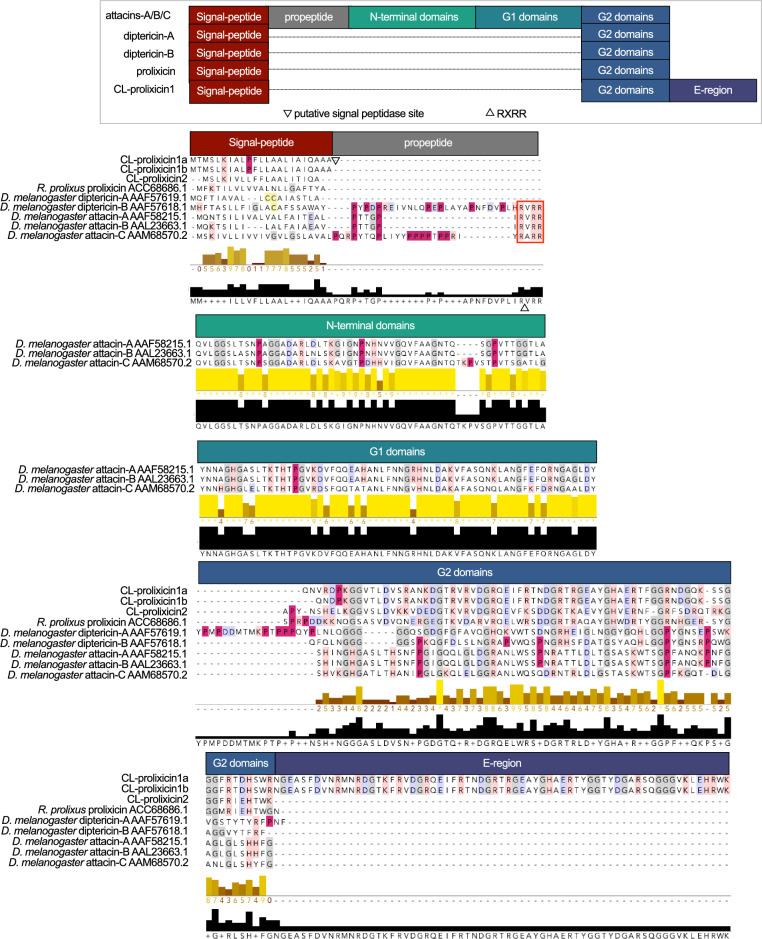
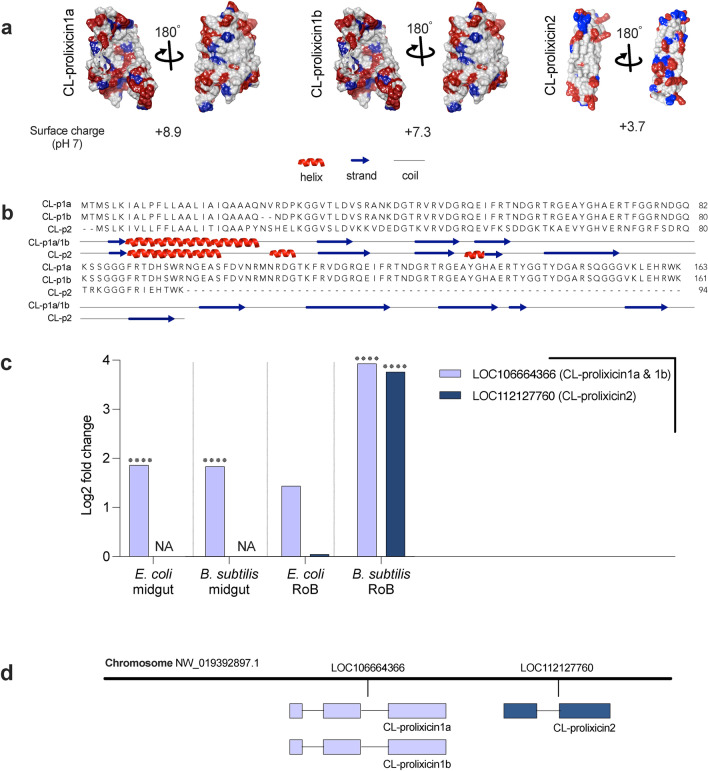
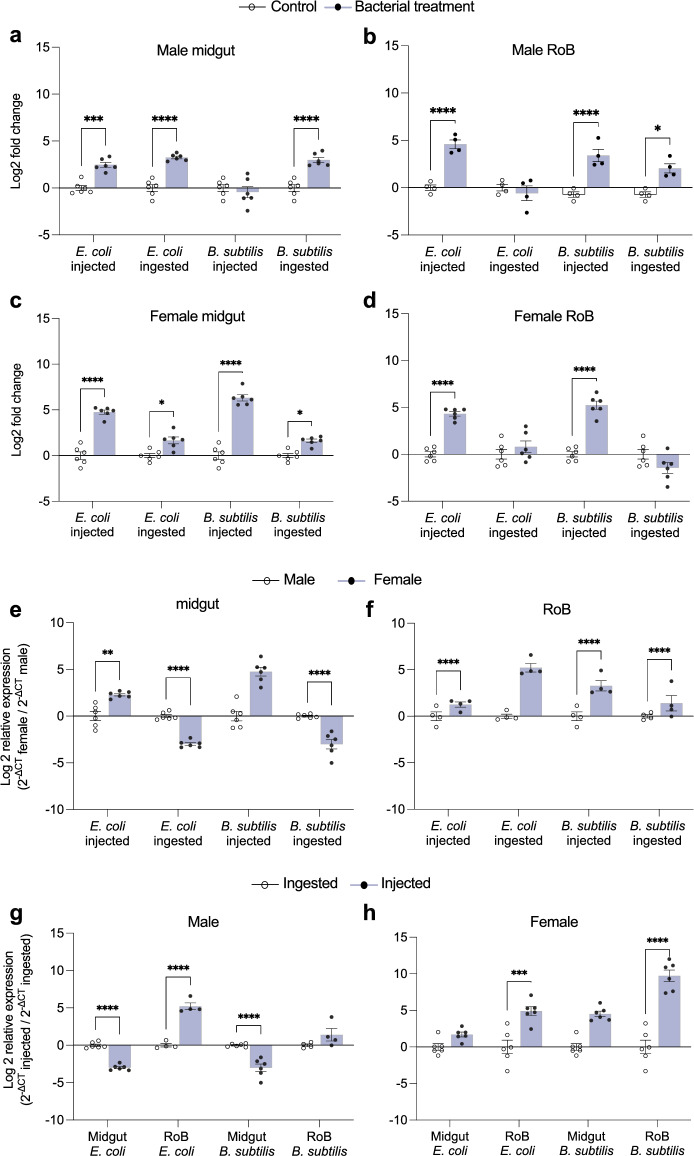
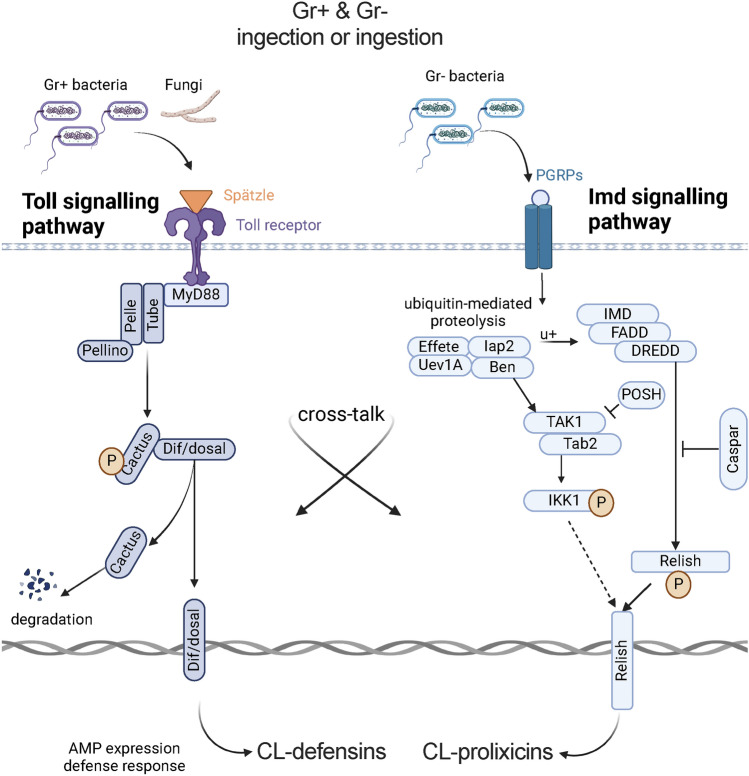
The hematophagous common bed bug, Cimex lectularius, is not known to transmit human pathogens outside laboratory settings, having evolved various immune defense mechanisms including the expression of antimicrobial peptides (AMPs). We unveil three novel prolixicin AMPs in bed bugs, exhibiting strong homology to the prolixicin of kissing bugs, Rhodnius prolixus, and to diptericin/attacin AMPs. We demonstrate for the first time sex-specific and immune mode-specific upregulation of these prolixicins in immune organs, the midgut and rest of body, following injection and ingestion of Gr+ (Bacillus subtilis) and Gr- (Escherichia coli) bacteria. Synthetic CL-prolixicin2 significantly inhibited growth of E. coli strains and killed or impeded Trypanosoma cruzi, the Chagas disease agent. Our findings suggest that prolixicins are regulated by both IMD and Toll immune pathways, supporting cross-talk and blurred functional differentiation between major immune pathways. The efficacy of CL-prolixicin2 against T. cruzi underscores the potential of AMPs in Chagas disease management.
Keywords: Trypanosoma cruzi; Bed bugs; Chagas disease; Glycine-rich antimicrobial peptides; Prolixicin; Toll and IMD humoral innate immunity.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
