

This is a preprint.
Targeting of SUMOylation leads to cBAF complex stabilization and disruption of the SS18::SSX transcriptome in Synovial Sarcoma
- PMID: 38883782
- PMCID: PMC11177989
- DOI: 10.21203/rs.3.rs-4362092/v1
Targeting of SUMOylation leads to cBAF complex stabilization and disruption of the SS18::SSX transcriptome in Synovial Sarcoma
Update in
-
Targeting SUMOylation promotes cBAF complex stabilization and disruption of the SS18::SSX transcriptome in synovial sarcoma.Nat Commun. 2025 Nov 5;16(1):9761. doi: 10.1038/s41467-025-64665-8. Nat Commun. 2025. PMID: 41193430 Free PMC article.
Abstract
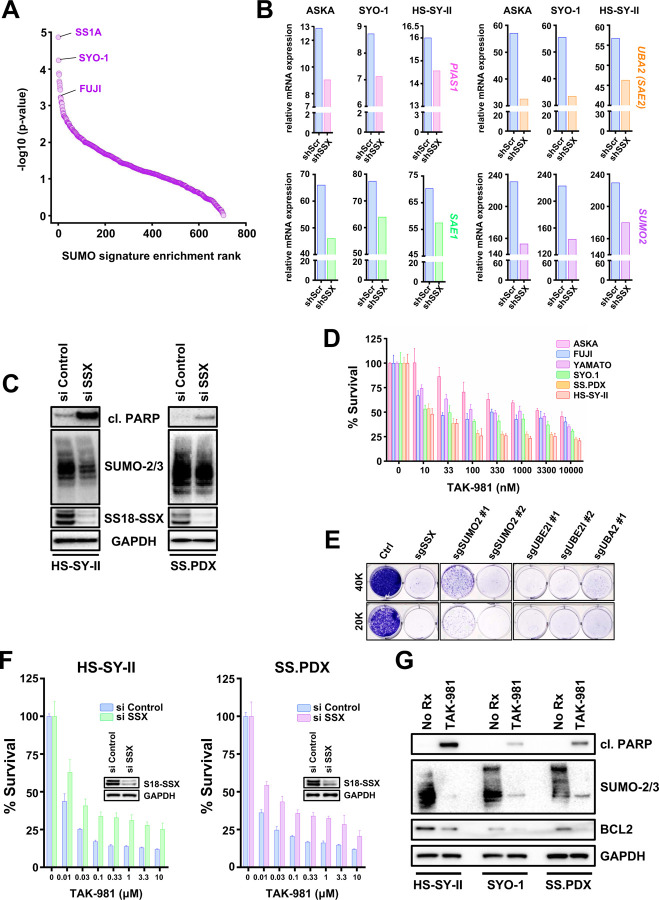
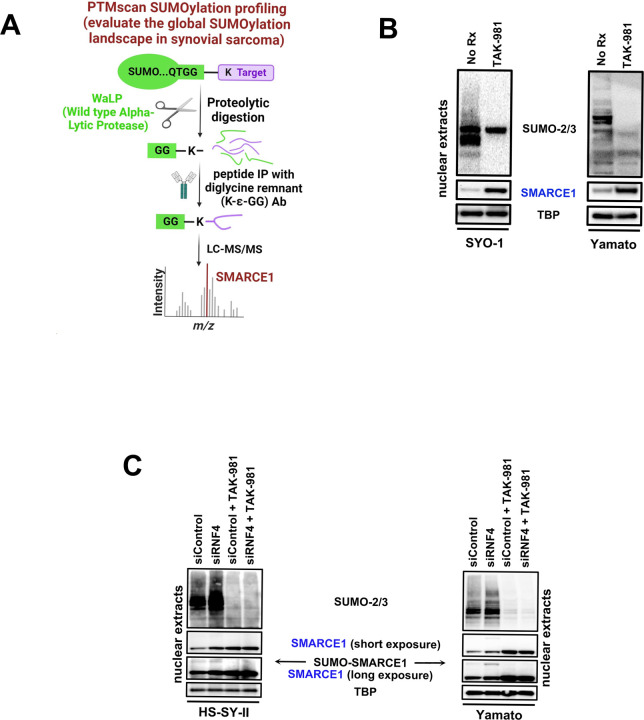
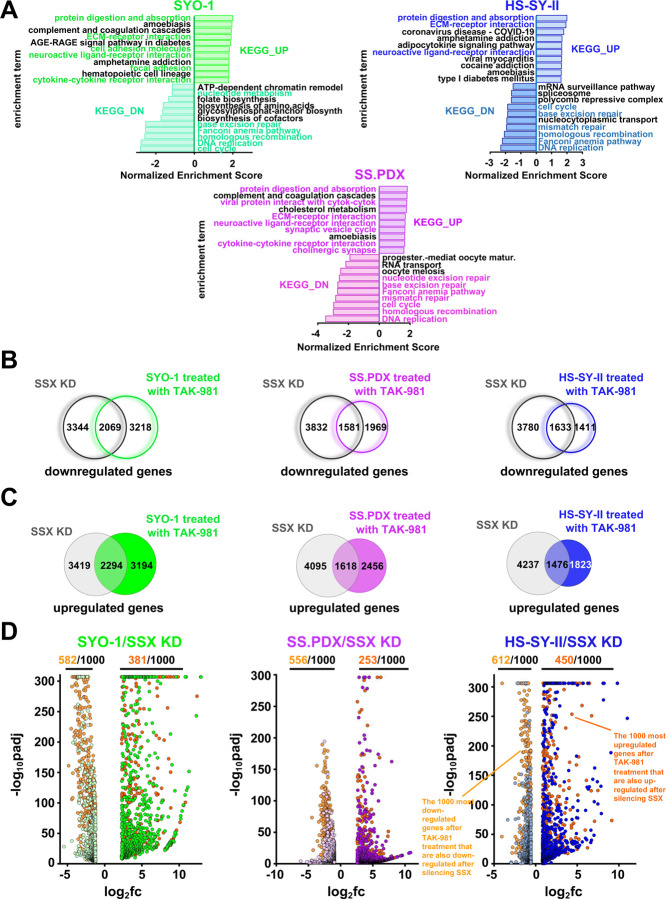
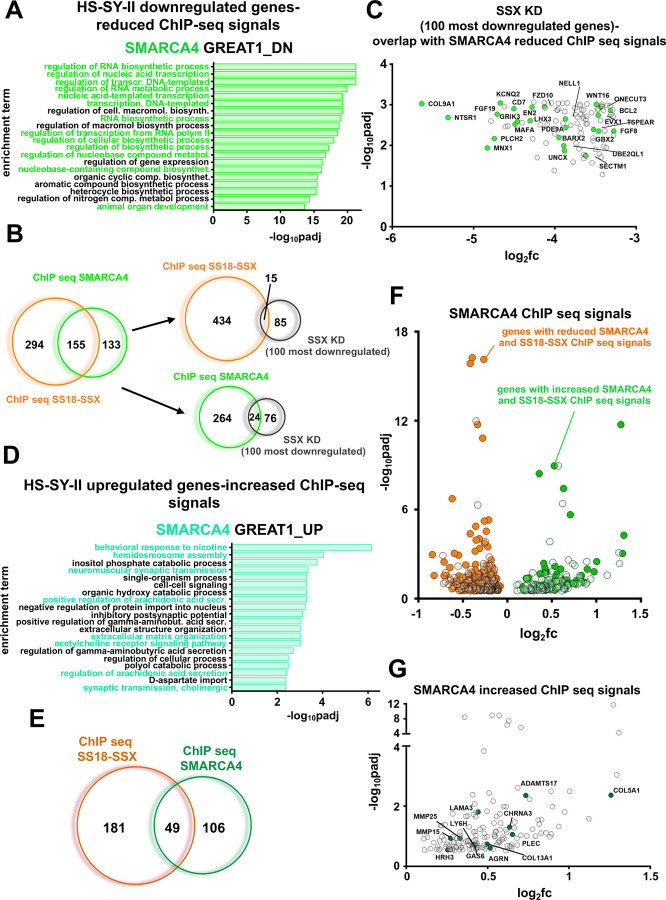
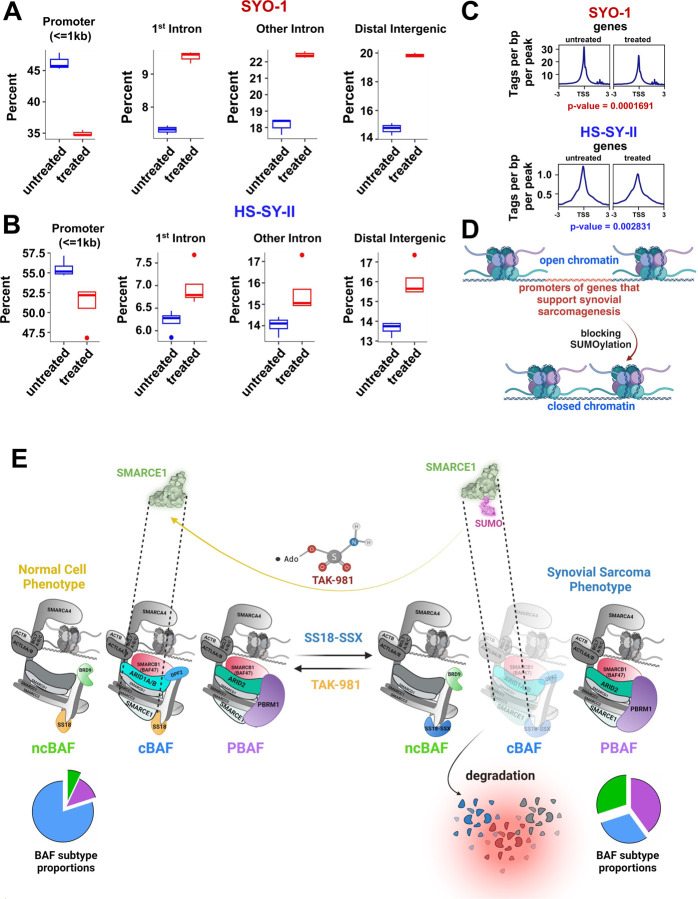
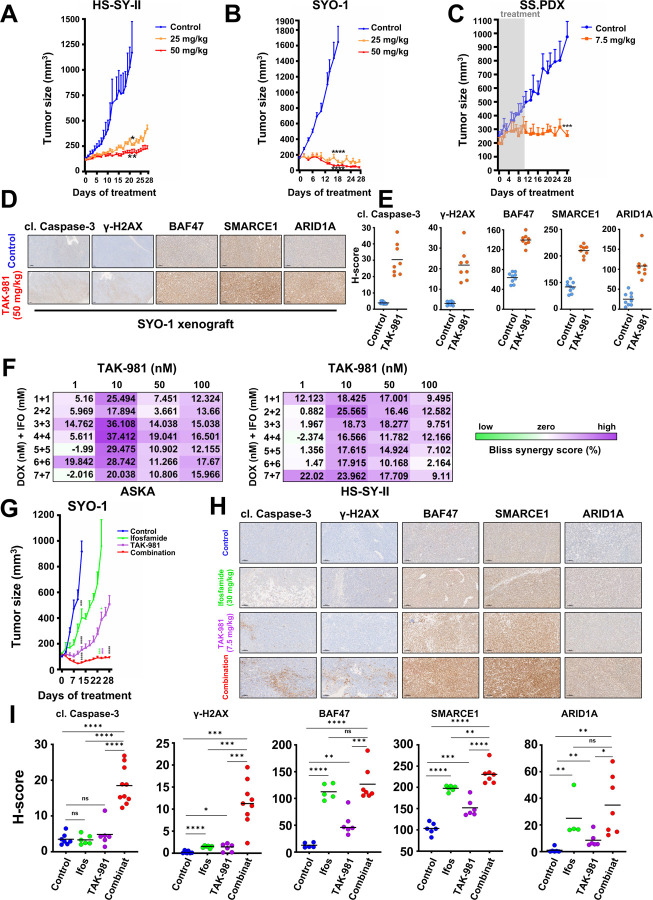
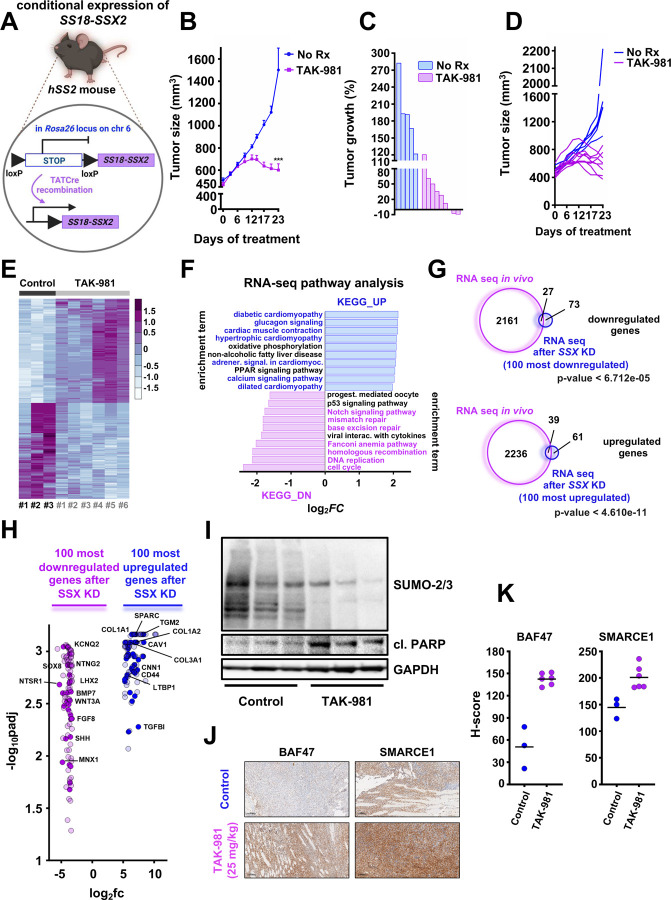
Synovial Sarcoma (SS) is driven by the SS18::SSX fusion oncoprotein and is ultimately refractory to therapeutic approaches. SS18::SSX alters ATP-dependent chromatin remodeling BAF (mammalian SWI/SNF) complexes, leading to the degradation of canonical (cBAF) complex and amplified presence of an SS18::SSX-containing non-canonical BAF (ncBAF or GBAF) that drives an SS-specific transcription program and tumorigenesis. We demonstrate that SS18::SSX activates the SUMOylation program and SSs are sensitive to the small molecule SAE1/2 inhibitor, TAK-981. Mechanistically, TAK-981 de-SUMOylates the cBAF subunit SMARCE1, stabilizing and restoring cBAF on chromatin, shifting away from SS18::SSX-ncBAF-driven transcription, associated with DNA damage and cell death and resulting in tumor inhibition across both human and mouse SS tumor models. TAK-981 synergized with cytotoxic chemotherapy through increased DNA damage, leading to tumor regression. Targeting the SUMOylation pathway in SS restores cBAF complexes and blocks the SS18::SSX-ncBAF transcriptome, identifying a therapeutic vulnerability in SS, positioning the in-clinic TAK-981 to treat SS.
Keywords: BAF complexes; DNA damage; SUMOylation; SWI/SNF; TAK-981; synovial sarcoma; targeted therapy.
Conflict of interest statement
Additional Declarations: There is NO Competing Interest. DECLARATION OF INTERESTS A.C.F. is a consultant and equity holder in Treeline Biosciences and has previously served as a scientific advisor for AbbVie and has received research funding from IDP Pharma. K.V. receives support from AstraZeneca. R.L. is an inventor of a provisional patent on targeted killing of EBV-positive cancer cells by CRISPR/dCas9-mediated EBV reactivation. S.B. is Consultant for Caris Lifescience and has received honoraria from SpringWorks for an educational lecture. K.V. receives support from AstraZeneca with no relationship to the present study. The authors declare that these listed activities have no relationship to the present study.
Figures

References
-
- Barrott J.J., Zhu J.F., Smith-Fry K., Susko A.M., Nollner D., Burrell L.D., Pozner A., Capecchi M.R., Yap J.T., Cannon-Albright L.A., et al. (2017). The Influential Role of BCL2 Family Members in Synovial Sarcomagenesis. Mol Cancer Res 15, 1733–1740. 10.1158/1541-7786.Mcr-17-0315. - DOI - PMC - PubMed
-
- Benjamin R.S., Legha S.S., Patel S.R., and Nicaise C. (1993). Single-agent ifosfamide studies in sarcomas of soft tissue and bone: the M.D. Anderson experience. Cancer Chemother Pharmacol 31 Suppl 2, S174–179. - PubMed
-
- Blay J.Y., von Mehren M., Jones R.L., Martin-Broto J., Stacchiotti S., Bauer S., Gelderblom H., Orbach D., Hindi N., Dei Tos A., and Nathenson M. (2023). Synovial sarcoma: characteristics, challenges, and evolving therapeutic strategies. ESMO Open 8, 101618. 10.1016/j.esmoop.2023.101618. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
