

Unveiling the crucial role of betaine: modulation of GABA homeostasis via SLC6A1 transporter (GAT1)
- PMID: 38884791
- PMCID: PMC11335192
- DOI: 10.1007/s00018-024-05309-w
Unveiling the crucial role of betaine: modulation of GABA homeostasis via SLC6A1 transporter (GAT1)
Abstract
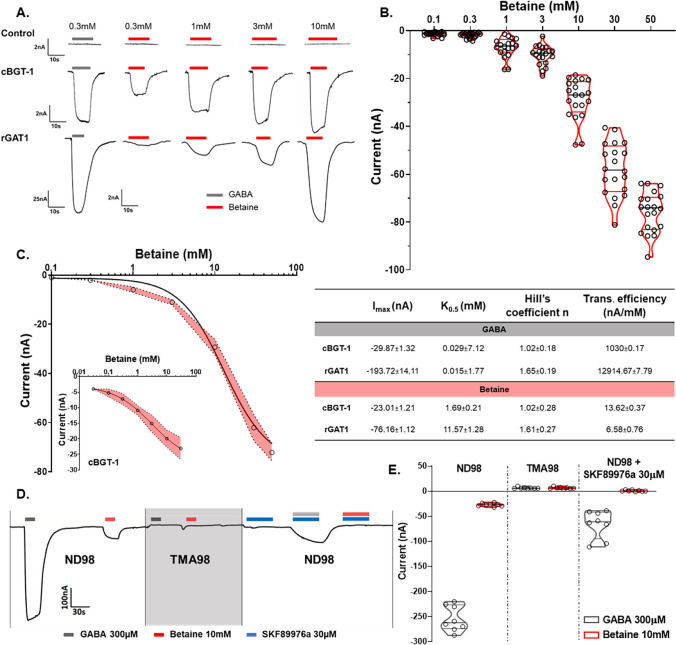
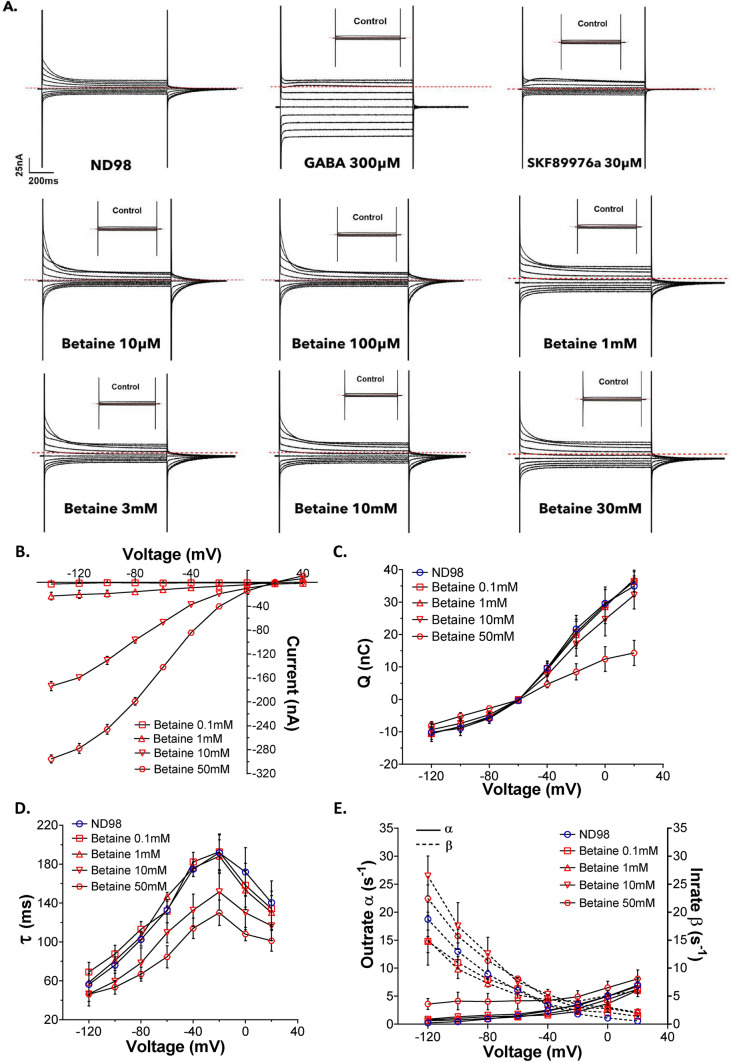
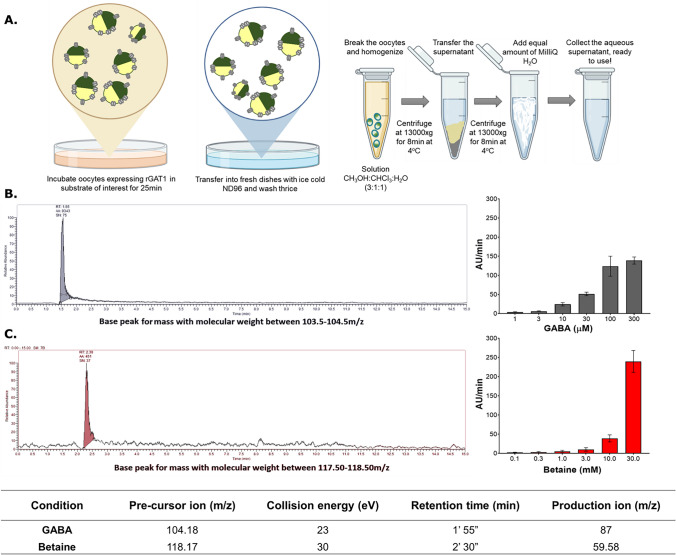
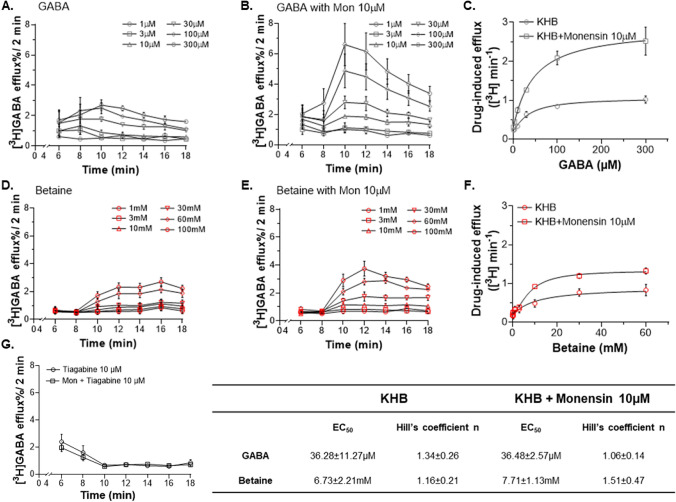
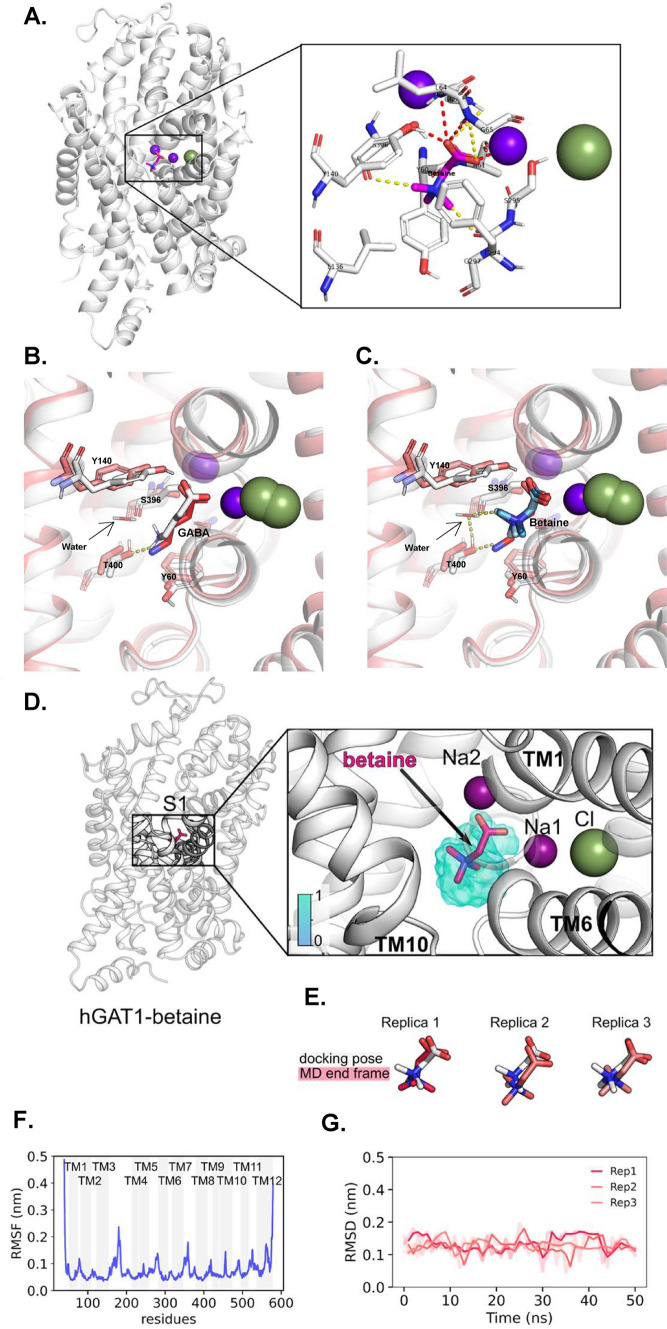
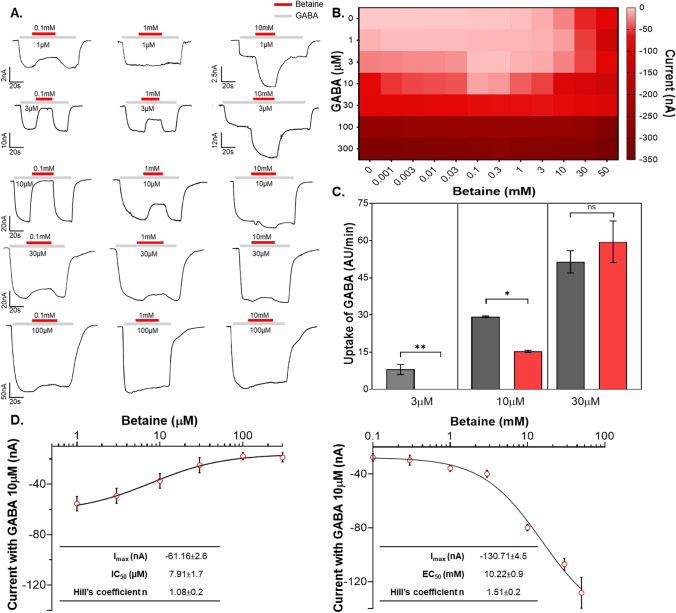
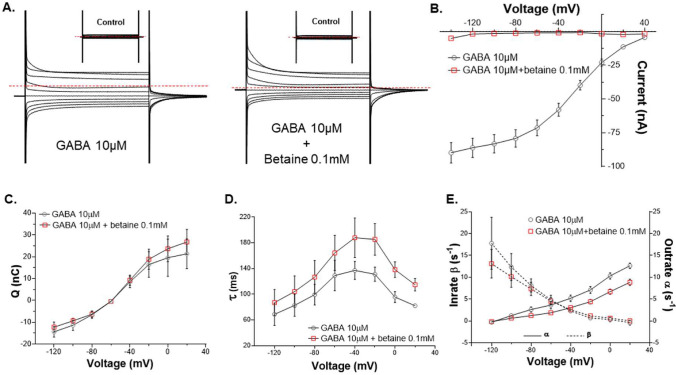
Betaine is an endogenous osmolyte that exhibits therapeutic potential by mitigating various neurological disorders. However, the underlying cellular and molecular mechanisms responsible for its neuroprotective effects remain puzzling.In this study, we describe a possible mechanism behind the positive impact of betaine in preserving neurons from excitotoxicity. Here we demonstrate that betaine at low concentration modulates the GABA uptake by GAT1 (slc6a1), the predominant GABA transporter in the central nervous system. This modulation occurs through the temporal inhibition of the transporter, wherein prolonged occupancy by betaine impedes the swift transition of the transporter to the inward conformation. Importantly, the modulatory effect of betaine on GAT1 is reversible, as the blocking of GAT1 disappears with increased extracellular GABA. Using electrophysiology, mass spectroscopy, radiolabelled cellular assay, and molecular dynamics simulation we demonstrate that betaine has a dual role in GAT1: at mM concentration acts as a slow substrate, and at µM as a temporal blocker of GABA, when it is below its K0.5. Given this unique modulatory characteristic and lack of any harmful side effects, betaine emerges as a promising neuromodulator of the inhibitory pathways improving GABA homeostasis via GAT1, thereby conferring neuroprotection against excitotoxicity.
Keywords: Betaine; E/I balance; GABA; GABA transporters; Neurological disorders; SLC6a1; Temporal inhibition.
© 2024. The Author(s).
Conflict of interest statement
AB and RZ were employed by Nanion technologies GmbH. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
