

FDXR variants cause adrenal insufficiency and atypical sexual development
- PMID: 38885337
- PMCID: PMC11383170
- DOI: 10.1172/jci.insight.179071
FDXR variants cause adrenal insufficiency and atypical sexual development
Abstract
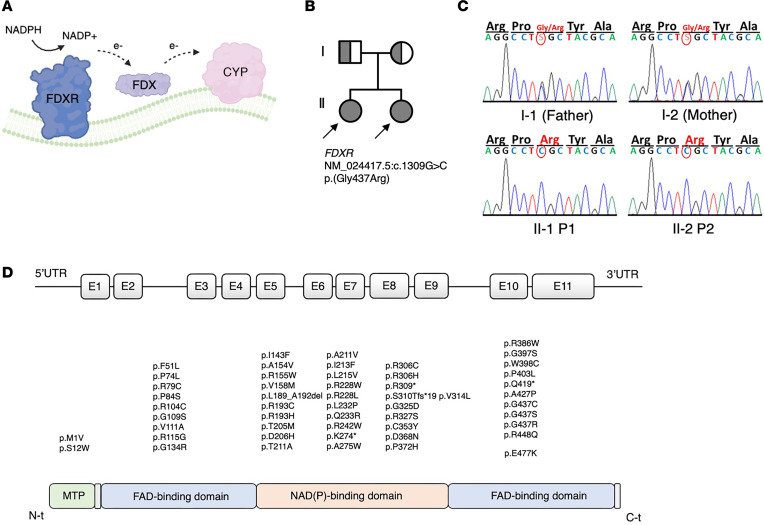
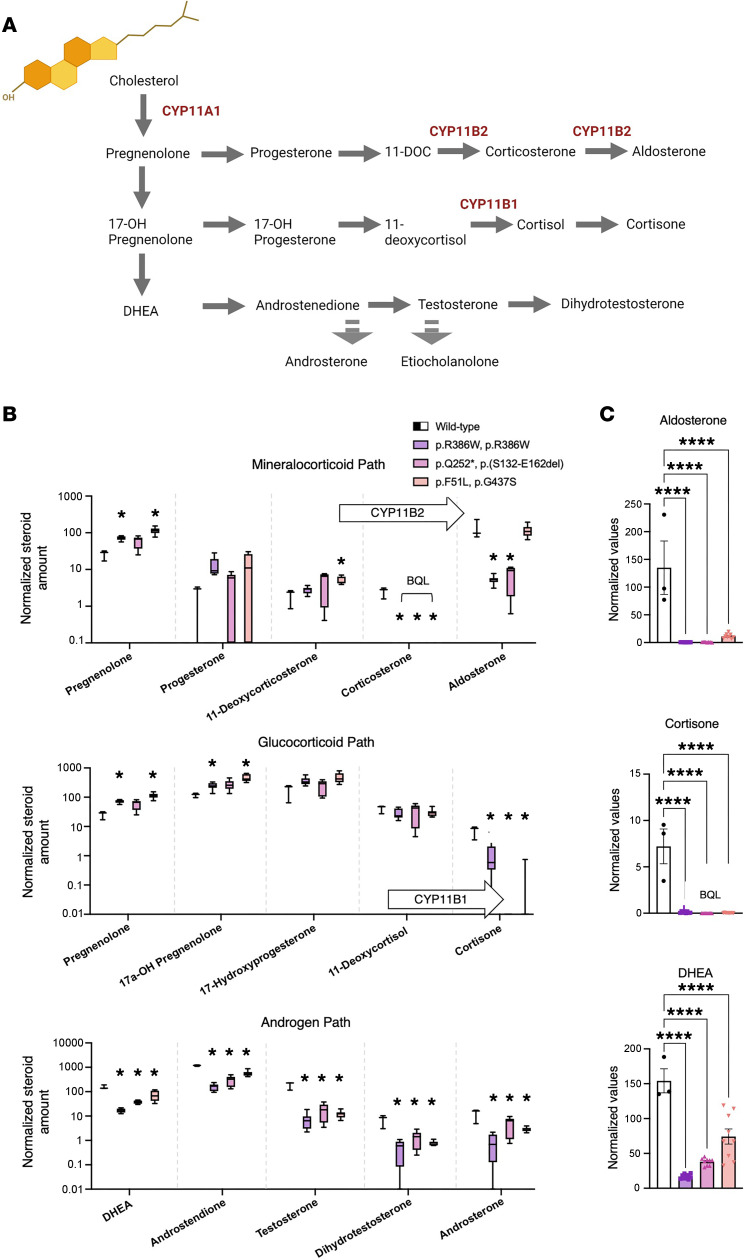
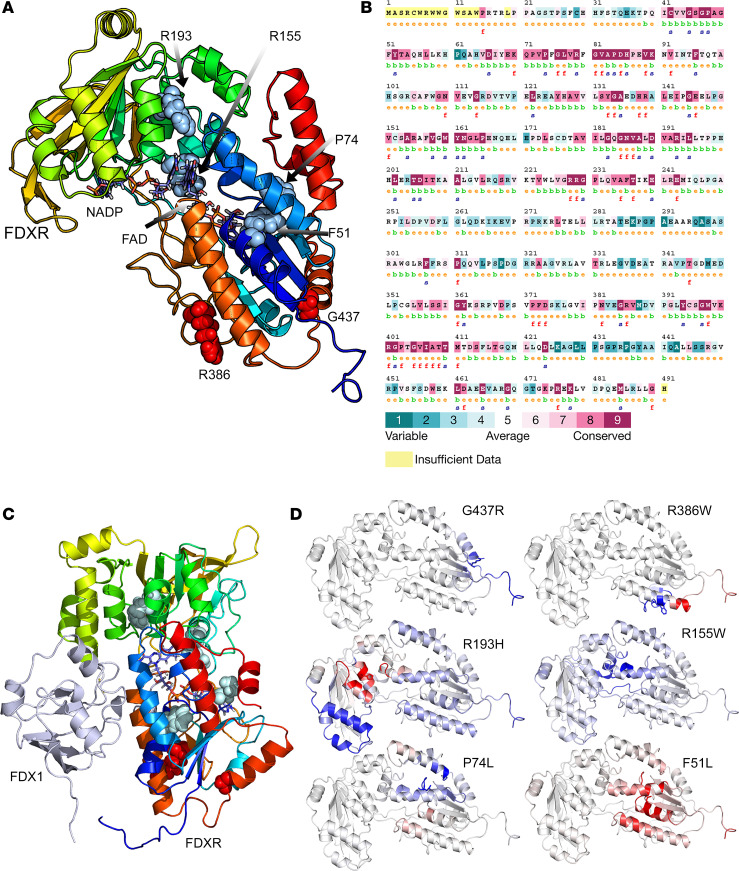
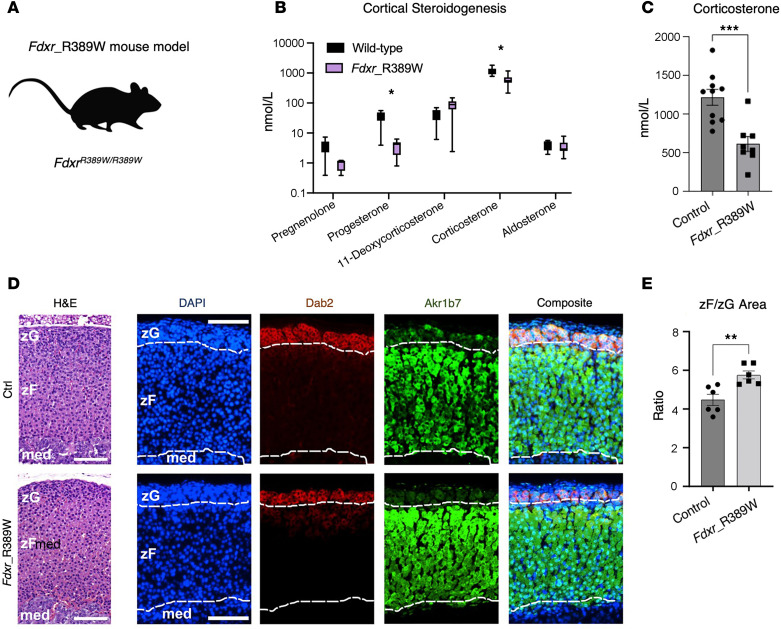
Genetic defects affecting steroid biosynthesis cause cortisol deficiency and differences of sex development; among these defects are recessive mutations in the steroidogenic enzymes CYP11A1 and CYP11B, whose function is supported by reducing equivalents donated by ferredoxin reductase (FDXR) and ferredoxin. So far, mutations in the mitochondrial flavoprotein FDXR have been associated with a progressive neuropathic mitochondriopathy named FDXR-related mitochondriopathy (FRM), but cortisol insufficiency has not been documented. However, patients with FRM often experience worsening or demise following stress associated with infections. We investigated 2 female patients with FRM carrying the potentially novel homozygous FDXR mutation p.G437R with ambiguous genitalia at birth and sudden death in the first year of life; they presented with cortisol deficiency and androgen excess compatible with 11-hydroxylase deficiency. In addition, steroidogenic FDXR-variant cell lines reprogrammed from 3 patients with FRM fibroblasts displayed deficient mineralocorticoid and glucocorticoid production. Finally, Fdxr-mutant mice allelic to the severe p.R386W human variant showed reduced progesterone and corticosterone production. Therefore, our comprehensive studies show that human FDXR variants may cause compensated but possibly life-threatening adrenocortical insufficiency in stress by affecting adrenal glucocorticoid and mineralocorticoid synthesis through direct enzyme inhibition, most likely in combination with disturbed mitochondrial redox balance.
Keywords: Endocrinology; Genetic diseases; Genetics; Mitochondria; Molecular genetics.
Figures
References
-
- Miller WL, et al. The Adrenal Cortex and Its Disorders. In: Sperling MA, ed. Sperling Pediatric Endocrinology, 5th Edition. Elsevier; 2020:425–490.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
