

The Mechanics of Building Functional Organs
- PMID: 38886066
- PMCID: PMC7616527
- DOI: 10.1101/cshperspect.a041520
The Mechanics of Building Functional Organs
Abstract
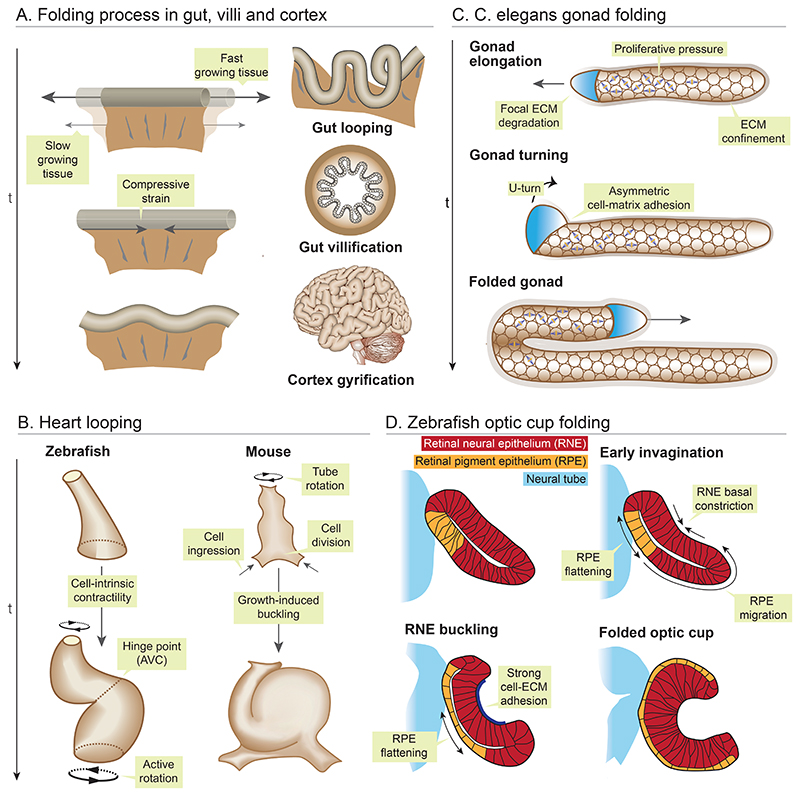
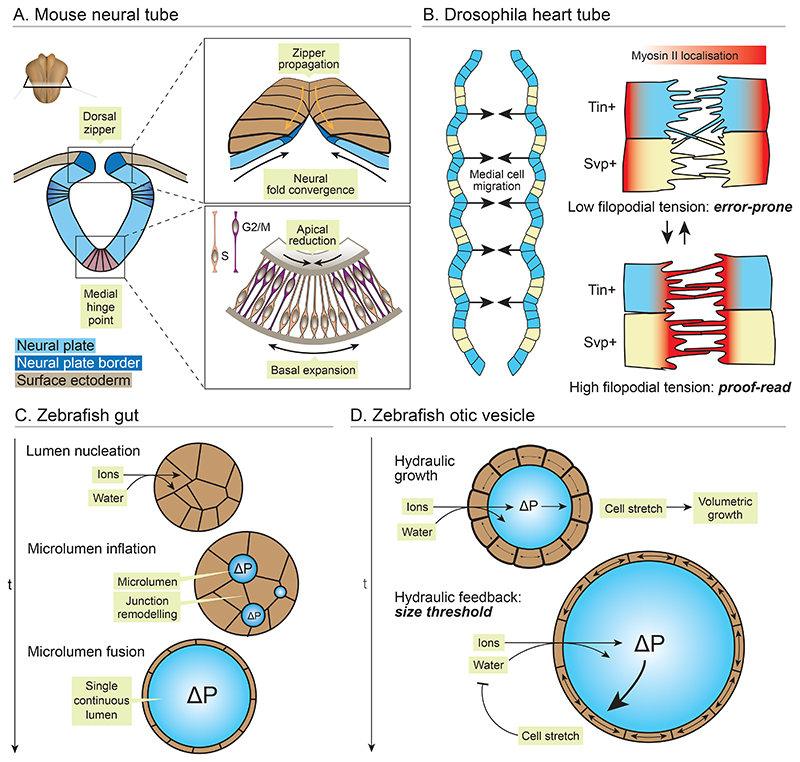
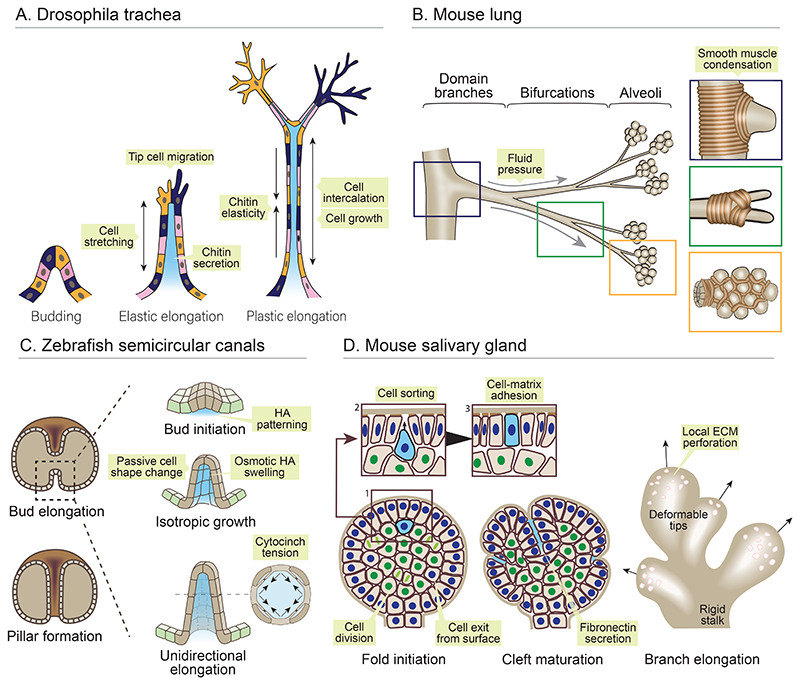
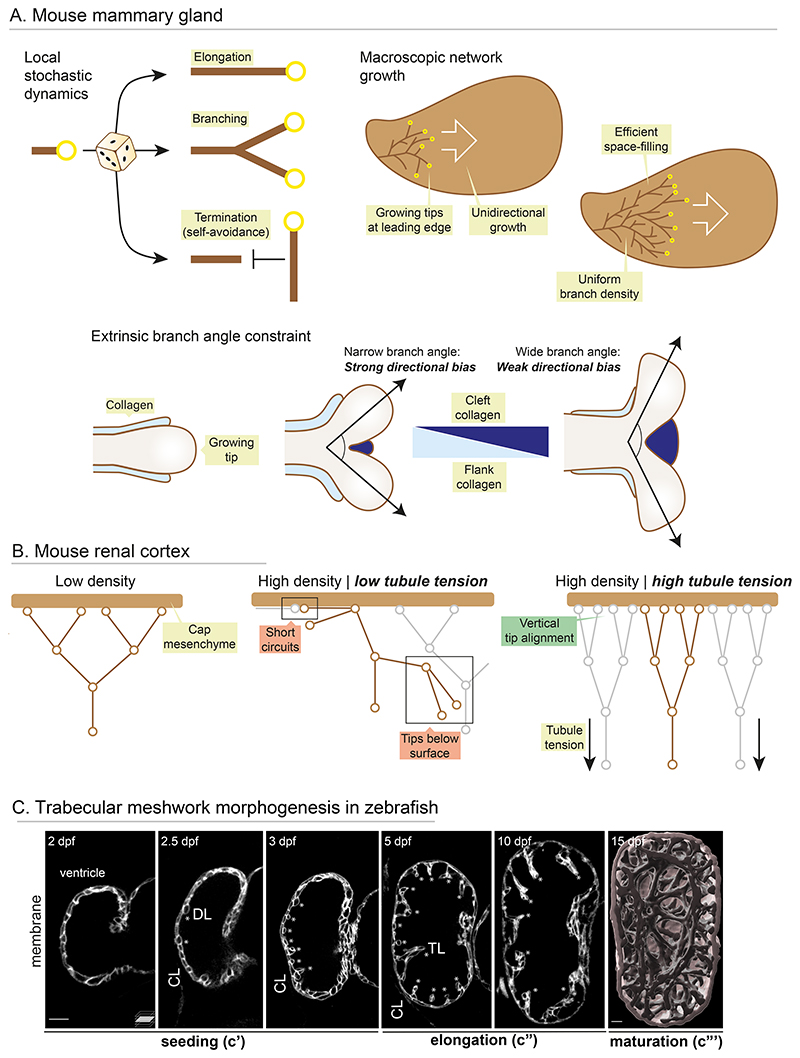
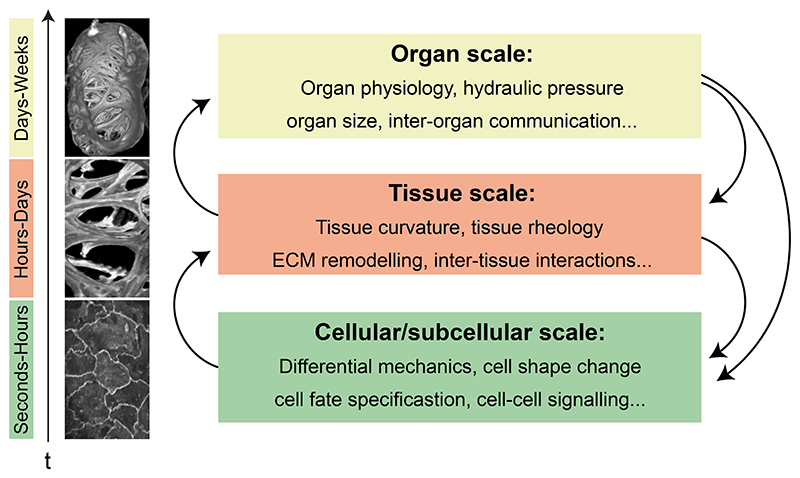
Organ morphogenesis is multifaceted, multiscale, and fundamentally a robust process. Despite the complex and dynamic nature of embryonic development, organs are built with reproducible size, shape, and function, allowing them to support organismal growth and life. This striking reproducibility of tissue form exists because morphogenesis is not entirely hardwired. Instead, it is an emergent product of mechanochemical information flow, operating across spatial and temporal scales-from local cellular deformations to organ-scale form and function, and back. In this review, we address the mechanical basis of organ morphogenesis, as understood by observations and experiments in living embryos. To this end, we discuss how mechanical information controls the emergence of a highly conserved set of structural motifs that shape organ architectures across the animal kingdom: folds and loops, tubes and lumens, buds, branches, and networks. Moving forward, we advocate for a holistic conceptual framework for the study of organ morphogenesis, which rests on an interdisciplinary toolkit and brings the embryo center stage.
Copyright © 2025 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources