

RANKL, but Not R-Spondins, Is Involved in Vascular Smooth Muscle Cell Calcification through LGR4 Interaction
- PMID: 38891922
- PMCID: PMC11172097
- DOI: 10.3390/ijms25115735
RANKL, but Not R-Spondins, Is Involved in Vascular Smooth Muscle Cell Calcification through LGR4 Interaction
Abstract
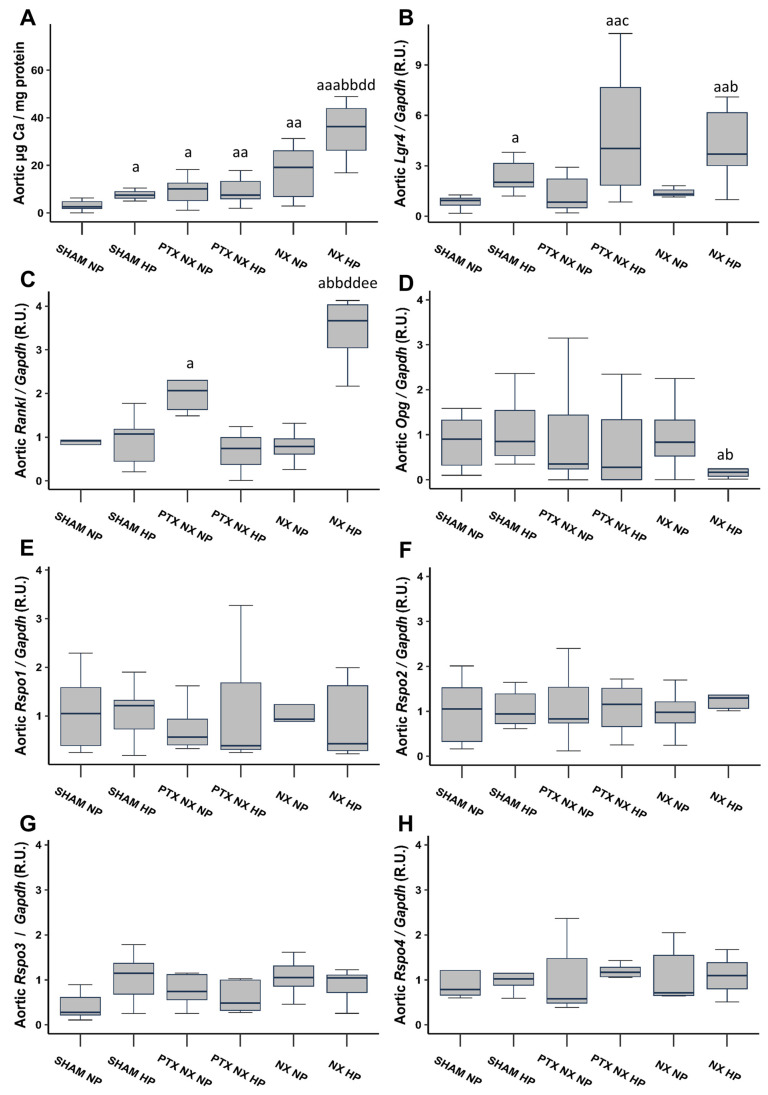
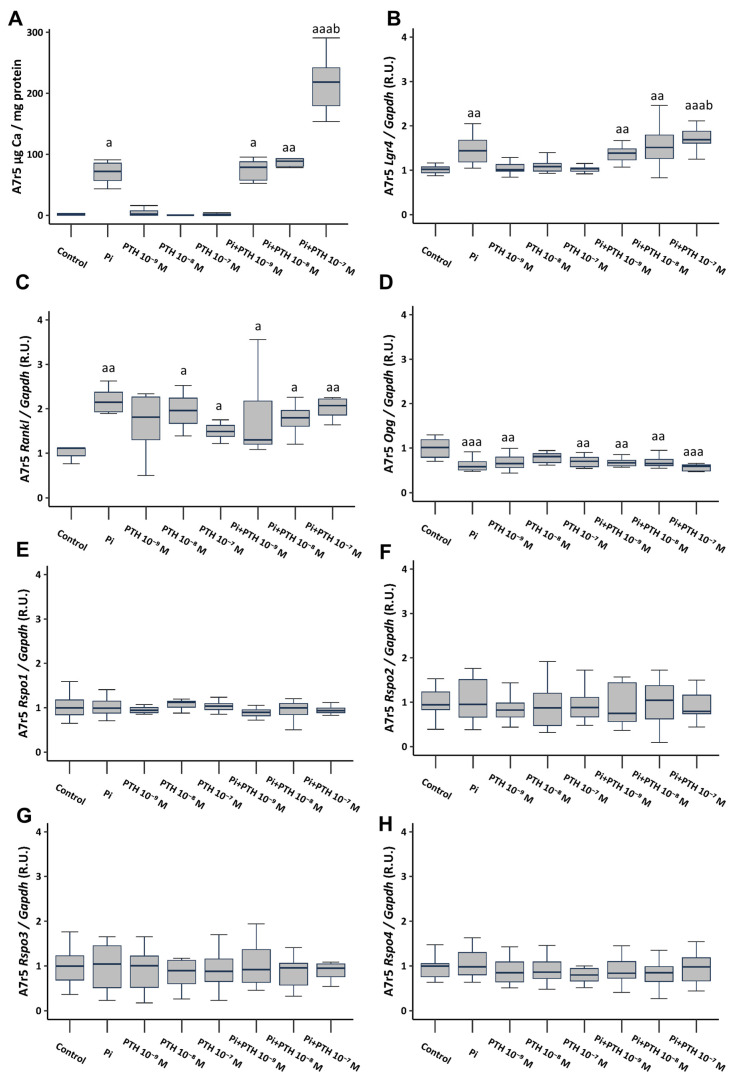
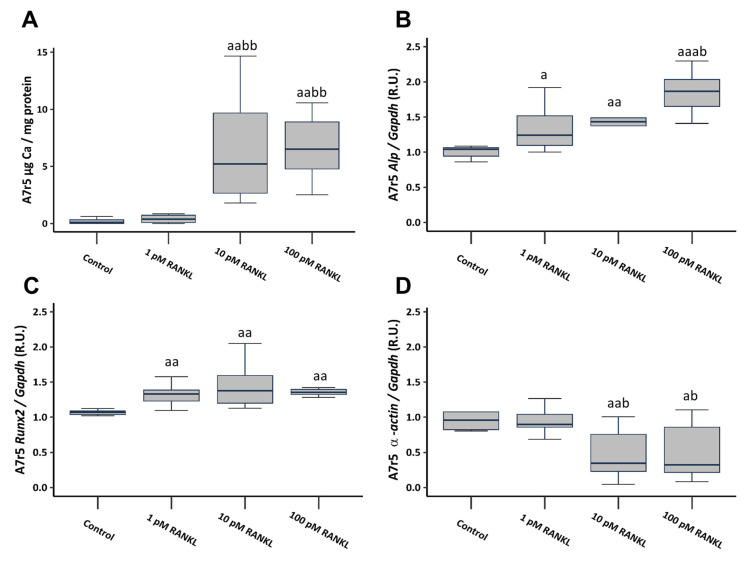
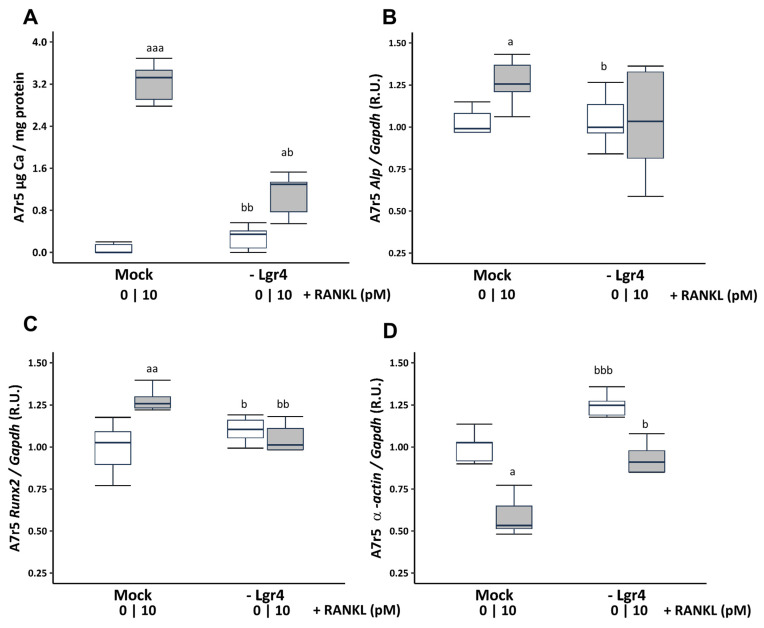
Vascular calcification has a global health impact that is closely linked to bone loss. The Receptor Activator of Nuclear Factor Kappa B (RANK)/RANK ligand (RANKL)/osteoprotegerin (OPG) system, fundamental for bone metabolism, also plays an important role in vascular calcification. The Leucine-rich repeat-containing G-protein-coupled receptor 4 (LGR4), a novel receptor for RANKL, regulates bone remodeling, and it appears to be involved in vascular calcification. Besides RANKL, LGR4 interacts with R-spondins (RSPOs), which are known for their roles in bone but are less understood in vascular calcification. Studies were conducted in rats with chronic renal failure fed normal or high phosphorus diets for 18 weeks, with and without control of circulating parathormone (PTH) levels, resulting in different degrees of aortic calcification. Additionally, vascular smooth muscle cells (VSMCs) were cultured under non-calcifying (1 mM phosphate) and calcifying (3 mM phosphate) media with different concentrations of PTH. To explore the role of RANKL in VSMC calcification, increasing concentrations of soluble RANKL were added to non-calcifying and calcifying media. The effects mediated by RANKL binding to its receptor LGR4 were investigated by silencing the LGR4 receptor in VSMCs. Furthermore, the gene expression of the RANK/RANKL/OPG system and the ligands of LGR4 was assessed in human epigastric arteries obtained from kidney transplant recipients with calcification scores (Kauppila Index). Increased aortic calcium in rats coincided with elevated systolic blood pressure, upregulated Lgr4 and Rankl gene expression, downregulated Opg gene expression, and higher serum RANKL/OPG ratio without changes in Rspos gene expression. Elevated phosphate in vitro increased calcium content and expression of Rankl and Lgr4 while reducing Opg. Elevated PTH in the presence of high phosphate exacerbated the increase in calcium content. No changes in Rspos were observed under the conditions employed. The addition of soluble RANKL to VSMCs induced genotypic differentiation and calcification, partly prevented by LGR4 silencing. In the epigastric arteries of individuals presenting vascular calcification, the gene expression of RANKL was higher. While RSPOs show minimal impact on VSMC calcification, RANKL, interacting with LGR4, drives osteogenic differentiation in VSMCs, unveiling a novel mechanism beyond RANKL-RANK binding.
Keywords: LGR4; R-spondins; RANKL; phosphorus; vascular calcification.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
The receptor activator of nuclear factor κΒ ligand receptor leucine-rich repeat-containing G-protein-coupled receptor 4 contributes to parathyroid hormone-induced vascular calcification.Nephrol Dial Transplant. 2021 Mar 29;36(4):618-631. doi: 10.1093/ndt/gfaa290. Nephrol Dial Transplant. 2021. PMID: 33367746
-
Increased calcification in osteoprotegerin-deficient smooth muscle cells: Dependence on receptor activator of NF-κB ligand and interleukin 6.J Vasc Res. 2014;51(2):118-31. doi: 10.1159/000358920. Epub 2014 Mar 7. J Vasc Res. 2014. PMID: 24642764 Free PMC article.
-
RANKL increases vascular smooth muscle cell calcification through a RANK-BMP4-dependent pathway.Circ Res. 2009 May 8;104(9):1041-8. doi: 10.1161/CIRCRESAHA.108.189001. Epub 2009 Mar 26. Circ Res. 2009. PMID: 19325147
-
Signaling pathways involved in vascular smooth muscle cell calcification during hyperphosphatemia.Cell Mol Life Sci. 2019 Jun;76(11):2077-2091. doi: 10.1007/s00018-019-03054-z. Epub 2019 Mar 18. Cell Mol Life Sci. 2019. PMID: 30887097 Free PMC article. Review.
-
Role of the RANK/RANKL/OPG and Wnt/β-Catenin Systems in CKD Bone and Cardiovascular Disorders.Calcif Tissue Int. 2021 Apr;108(4):439-451. doi: 10.1007/s00223-020-00803-2. Epub 2021 Feb 13. Calcif Tissue Int. 2021. PMID: 33586001 Review.
References
-
- Almaden Y., Hernandez A., Torregrosa V., Canalejo A., Sabate L., Fernandez Cruz L., Campistol J.M., Torres A., Rodriguez M. High phosphate level directly stimulates parathyroid hormone secretion and synthesis by human parathyroid tissue in vitro. J. Am. Soc. Nephrol. JASN. 1998;9:1845–1852. doi: 10.1681/ASN.V9101845. - DOI - PubMed
MeSH terms
Substances
Grants and funding
- CP23/00105; CP23/00058; PI20/00753, PI20/00633, PI22/00195, and PI23/00833 co-funded by the European Union/Instituto de Salud Carlos III (ISCIII)
- RD16/0009/0017/ISCIII Retic REDinREN
- RD21/0005/0019 and RD21/0005/0011/ISCIII Red de Investigación Cooperativa Orientada a Resultados en Salud RICORS2040, Next Generation EU; Recovery, Transformation and Resilience Plan
- GRUPIN14-028, IDI-2018-000152, IDI/2021/000080/Plan Estatal de I + D + I 2013-2016, Plan de Ciencia, Tecnología e Innovación 2013-2017 y 2018-2022 del Principado de Asturias
- Iñigo Alvarez de Toledo/Fundación Renal Iñigo Álvarez de Toledo (FRIAT)
LinkOut - more resources
Full Text Sources
