

This is a preprint.
Protein absorption in the zebrafish gut is regulated by interactions between lysosome rich enterocytes and the microbiome
- PMID: 38895310
- PMCID: PMC11185774
- DOI: 10.1101/2024.06.07.597998
Protein absorption in the zebrafish gut is regulated by interactions between lysosome rich enterocytes and the microbiome
Update in
-
Protein absorption in the zebrafish gut is regulated by interactions between lysosome rich enterocytes and the microbiome.Elife. 2025 Mar 13;13:RP100611. doi: 10.7554/eLife.100611. Elife. 2025. PMID: 40080061 Free PMC article.
Abstract
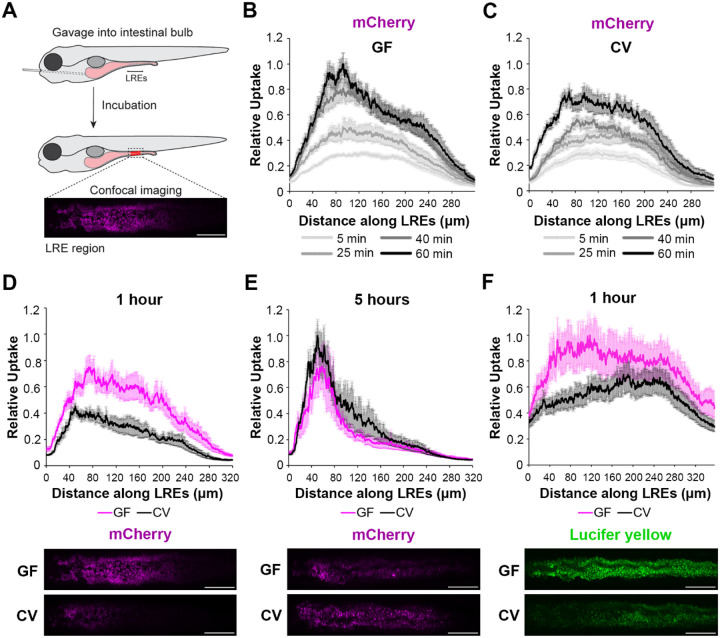
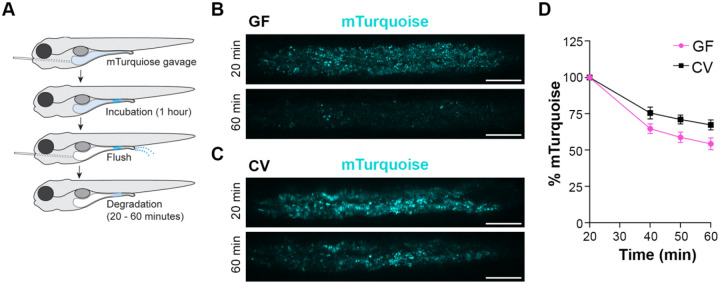
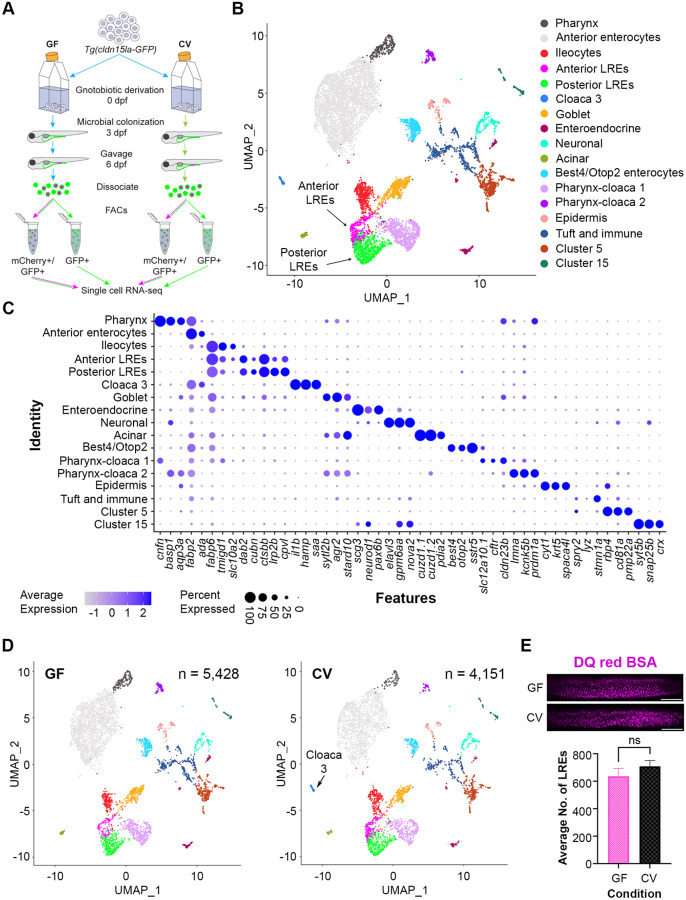
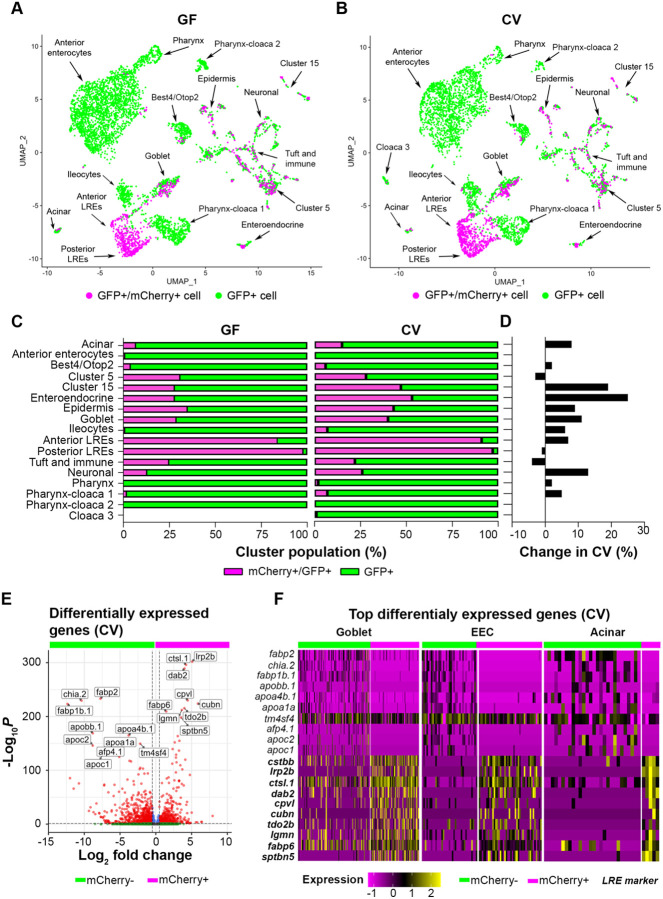
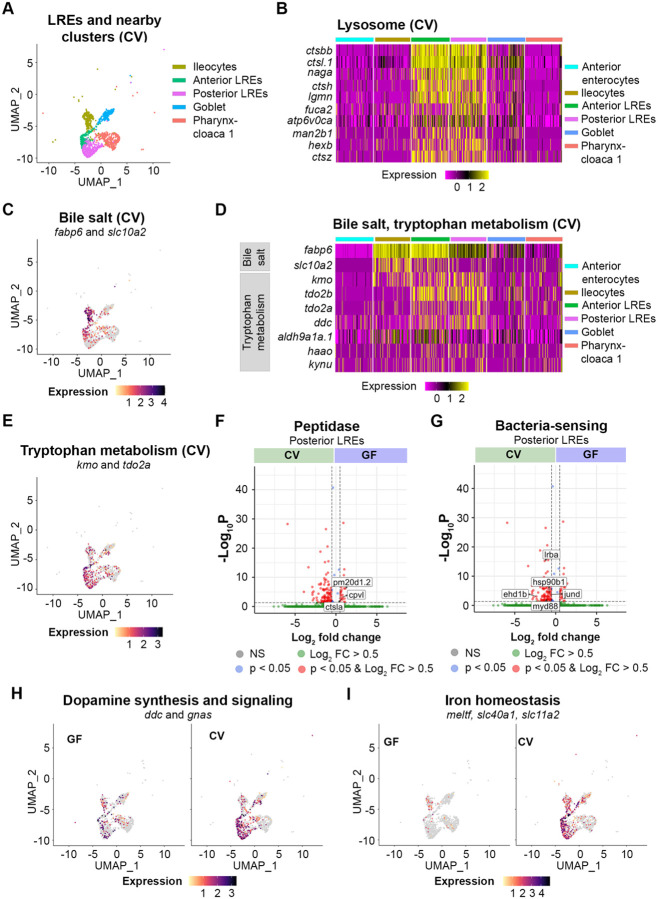
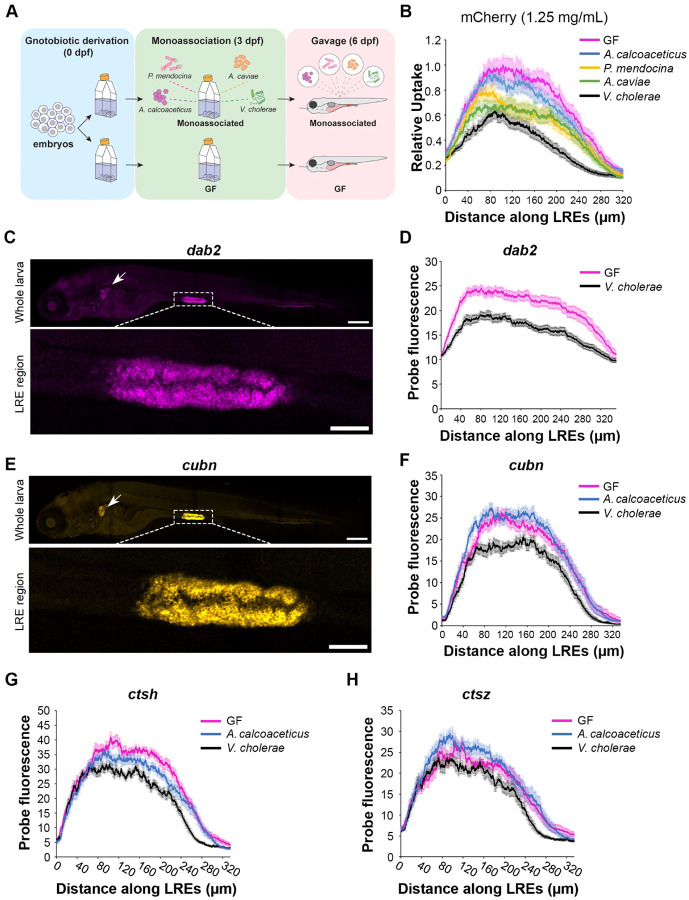
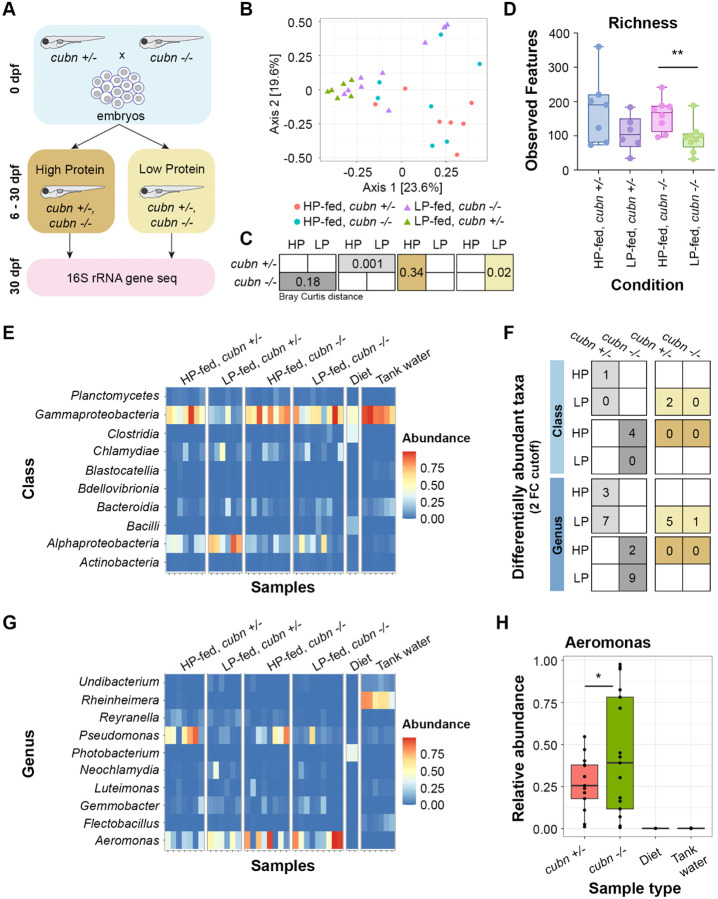
Dietary protein absorption in neonatal mammals and fishes relies on the function of a specialized and conserved population of highly absorptive lysosome rich enterocytes (LREs). The gut microbiome has been shown to enhance absorption of nutrients, such as lipids, by intestinal epithelial cells. However, whether protein absorption is also affected by the gut microbiome is poorly understood. Here, we investigate connections between protein absorption and microbes in the zebrafish gut. Using live microscopy-based quantitative assays, we find that microbes slow the pace of protein uptake and degradation in LREs. While microbes do not affect the number of absorbing LRE cells, microbes lower the expression of endocytic and protein digestion machinery in LREs. Using transgene assisted cell isolation and single cell RNA-sequencing, we characterize all intestinal cells that take up dietary protein. We find that microbes affect expression of bacteria-sensing and metabolic pathways in LREs, and that some secretory cell types also take up protein and share components of protein uptake and digestion machinery with LREs. Using custom-formulated diets, we investigated the influence of diet and LRE activity on the gut microbiome. Impaired protein uptake activity in LREs, along with a protein-deficient diet, alters the microbial community and leads to increased abundance of bacterial genera that have the capacity to reduce protein uptake in LREs. Together, these results reveal that diet-dependent reciprocal interactions between LREs and the gut microbiome regulate protein absorption.
Figures
References
-
- Ahuja Malini, Schwartz Daniella M., Tandon Mayank, Son Aran, Zeng Mei, Swaim William, Eckhaus Michael, et al. 2017. “Orai1-Mediated Antimicrobial Secretion from Pancreatic Acini Shapes the Gut Microbiome and Regulates Gut Innate Immunity.” Cell Metabolism 25 (3): 635–46. 10.1016/j.cmet.2017.02.007. - DOI - PMC - PubMed
-
- Bao Honghong, Wang Yi, Xiong Hanlin, Xia Yaoyao, Cui Zhifu, and Liu Lingbin. 2024. “Mechanism of Iron Ion Homeostasis in Intestinal Immunity and Gut Microbiota Remodeling.” International Journal of Molecular Sciences. Multidisciplinary Digital Publishing Institute (MDPI). 10.3390/ijms25020727. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources