

This is a preprint.
Evidence of RNA polymerase III recruitment and transcription at protein-coding gene promoters
- PMID: 38895345
- PMCID: PMC11185800
- DOI: 10.1101/2024.06.08.598009
Evidence of RNA polymerase III recruitment and transcription at protein-coding gene promoters
Update in
-
Evidence of RNA polymerase III recruitment and transcription at protein-coding gene promoters.Mol Cell. 2024 Nov 7;84(21):4111-4124.e5. doi: 10.1016/j.molcel.2024.09.019. Epub 2024 Oct 10. Mol Cell. 2024. PMID: 39393362 Free PMC article.
Abstract
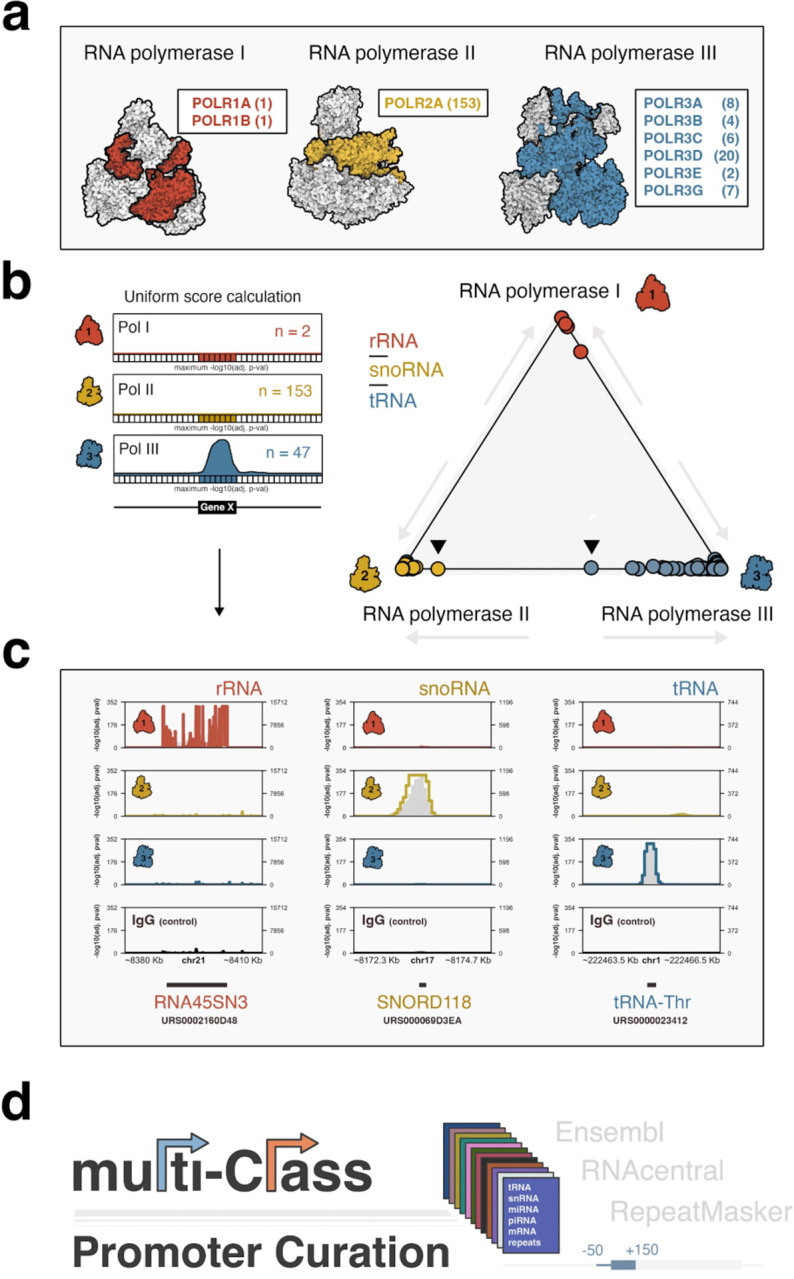
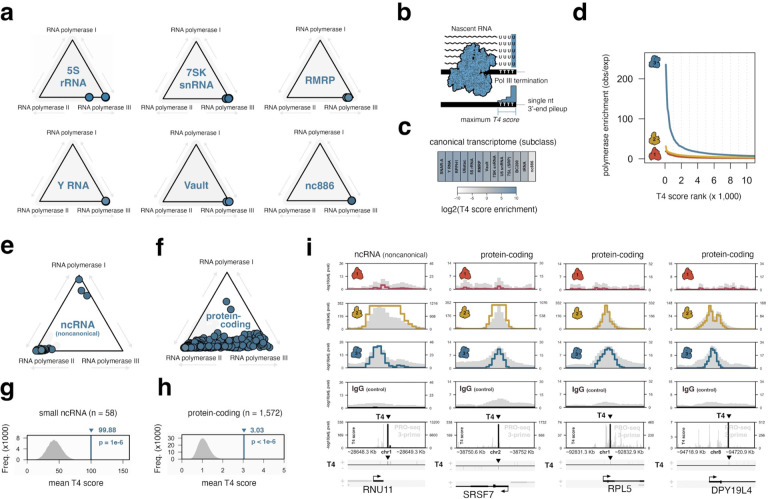
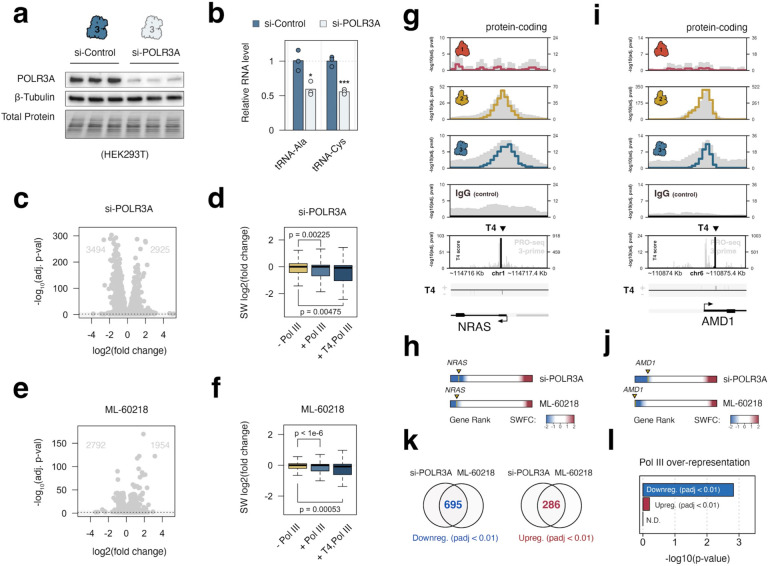
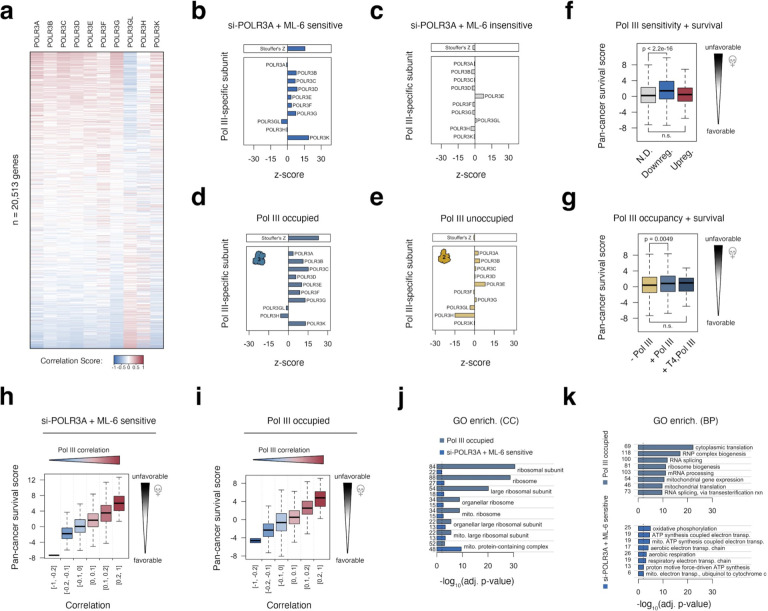
RNA polymerase (Pol) I, II, and III are most commonly described as having distinct roles in synthesizing ribosomal RNA (rRNA), messenger RNA (mRNA), and specific small noncoding (nc)RNAs, respectively. This delineation of transcriptional responsibilities is not definitive, however, as evidenced by instances of Pol II recruitment to genes conventionally transcribed by Pol III, including the co-transcription of RPPH1 - the catalytic RNA component of RNase P. A comprehensive understanding of the interplay between RNA polymerase complexes remains lacking, however, due to limited comparative analyses for all three enzymes. To address this gap, we applied a uniform framework for quantifying global Pol I, II, and III occupancies that integrates currently available human RNA polymerase ChIP-seq datasets. Occupancy maps are combined with a comprehensive multi-class promoter set that includes protein-coding genes, noncoding genes, and repetitive elements. While our genomic survey appropriately identifies recruitment of Pol I, II, and III to canonical target genes, we unexpectedly discover widespread recruitment of the Pol III machinery to promoters of specific protein-coding genes, supported by colocalization patterns observed for several Pol III-specific subunits. We show that Pol III-occupied Pol II promoters are enriched for small, nascent RNA reads terminating in a run of 4 Ts, a unique hallmark of Pol III transcription termination and evidence of active Pol III activity at these sites. Pol III disruption differentially modulates the expression of Pol III-occupied coding genes, which are functionally enriched for ribosomal proteins and genes broadly linked to unfavorable outcomes in cancer. Our map also identifies additional, currently unannotated genomic elements occupied by Pol III with clear signatures of nascent RNA species that are sensitive to disruption of La (SSB) - a Pol III-related RNA chaperone protein. These findings reshape our current understanding of the interplay between Pols II and III and identify potentially novel small ncRNAs with broad implications for gene regulatory paradigms and RNA biology.
Conflict of interest statement
Competing interest statement The authors declare no competing interests
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous