

Ubiquitination is involved in PKC-mediated degradation of cell surface Kv1.5 channels
- PMID: 38897569
- PMCID: PMC11301065
- DOI: 10.1016/j.jbc.2024.107483
Ubiquitination is involved in PKC-mediated degradation of cell surface Kv1.5 channels
Abstract
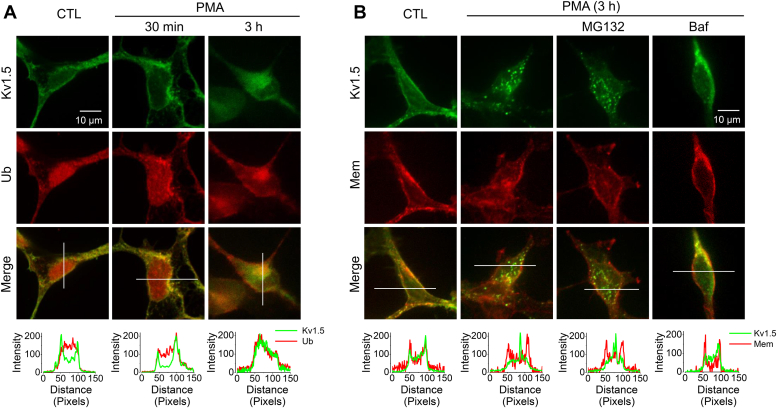
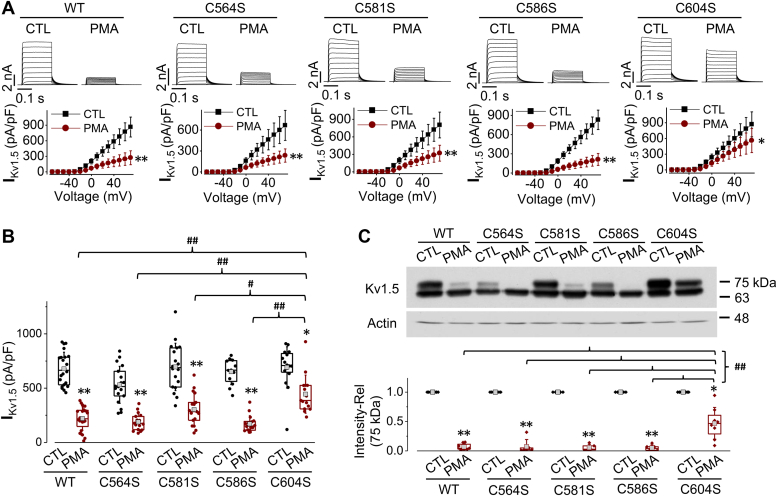
The voltage-gated Kv1.5 potassium channel, conducting the ultra-rapid delayed rectifier K+ current (IKur) in human cells, plays important roles in the repolarization of atrial action potentials and regulation of the vascular tone. We previously reported that activation of protein kinase C (PKC) by phorbol 12-myristate 13-acetate (PMA) induces endocytic degradation of cell-surface Kv1.5 channels, and a point mutation removing the phosphorylation site, T15A, in the N terminus of Kv1.5 abolished the PMA-effect. In the present study, using mutagenesis, patch clamp recording, Western blot analysis, and immunocytochemical staining, we demonstrate that ubiquitination is involved in the PMA-mediated degradation of mature Kv1.5 channels. Since the expression of the Kv1.4 channel is unaffected by PMA treatment, we swapped the N- and/or C-termini between Kv1.5 and Kv1.4. We found that the N-terminus alone did not but both N- and C-termini of Kv1.5 did confer PMA sensitivity to mature Kv1.4 channels, suggesting the involvement of Kv1.5 C-terminus in the channel ubiquitination. Removal of each of the potential ubiquitination residue Lysine at position 536, 565, and 591 by Arginine substitution (K536R, K565R, and K591R) had little effect, but removal of all three Lysine residues with Arginine substitution (3K-R) partially reduced PMA-mediated Kv1.5 degradation. Furthermore, removing the cysteine residue at position 604 by Serine substitution (C604S) drastically reduced PMA-induced channel degradation. Removal of the three Lysines and Cys604 with a quadruple mutation (3K-R/C604S) or a truncation mutation (Δ536) completely abolished the PKC activation-mediated degradation of Kv1.5 channels. These results provide mechanistic insight into PKC activation-mediated Kv1.5 degradation.
Keywords: patch clamp; protein kinase C; ubiquitination; voltage-gated potassium channel.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Albrecht B., Weber K., Pongs O. Characterization of a voltage-activated K-channel gene cluster on human chromosome 12p13. Recept Channels. 1995;3:213–220. - PubMed
-
- Fedida D., Wible B., Wang Z., Fermini B., Faust F., Nattel S., et al. Identity of a novel delayed rectifier current from human heart with a cloned K+ channel current. Circ. Res. 1993;73:210–216. - PubMed
-
- Wang Z., Fermini B., Nattel S. Sustained depolarization-induced outward current in human atrial myocytes: evidence for a novel delayed rectifier K+ current similar to Kv1.5 cloned channel currents. Circ. Res. 1993;73:1061–1076. - PubMed
-
- MacDonald P.E., Wheeler M.B. Voltage-dependent K+ channels in pancreatic beta cells: role, regulation and potential as therapeutic targets. Diabetologia. 2003;46:1046–1062. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
