

Single missense mutations in Vi capsule synthesis genes confer hypervirulence to Salmonella Typhi
- PMID: 38898034
- PMCID: PMC11187135
- DOI: 10.1038/s41467-024-49590-6
Single missense mutations in Vi capsule synthesis genes confer hypervirulence to Salmonella Typhi
Abstract
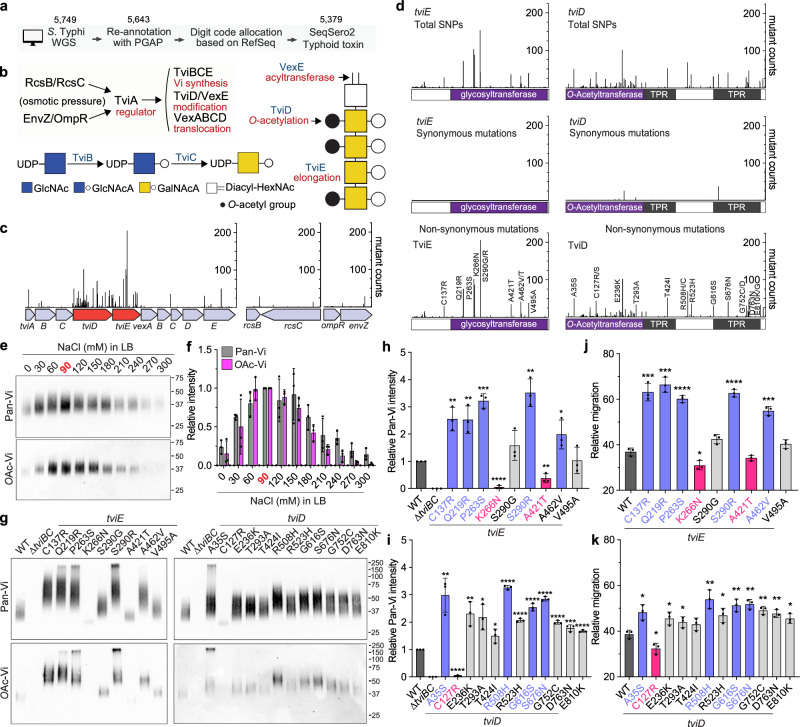
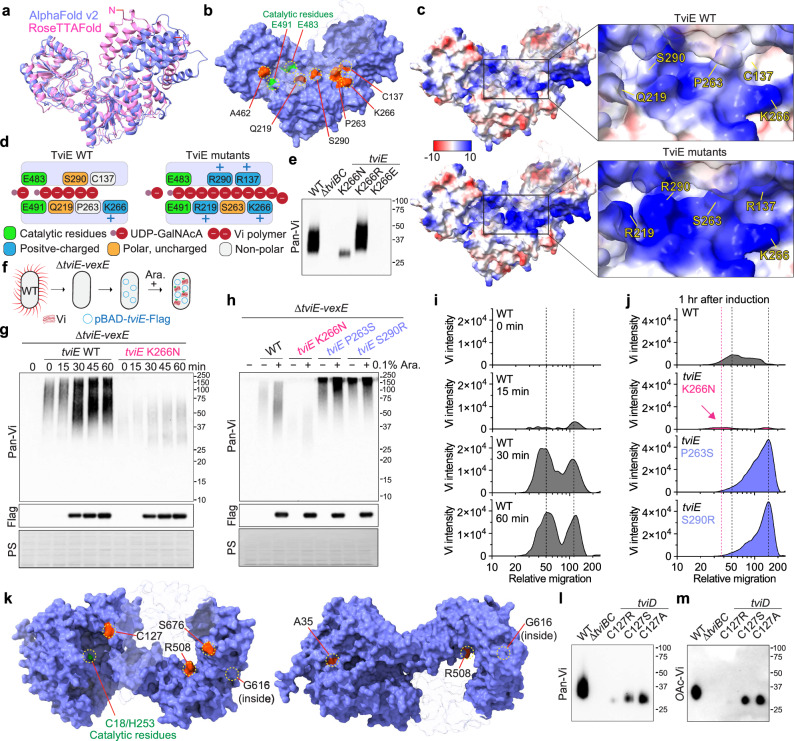
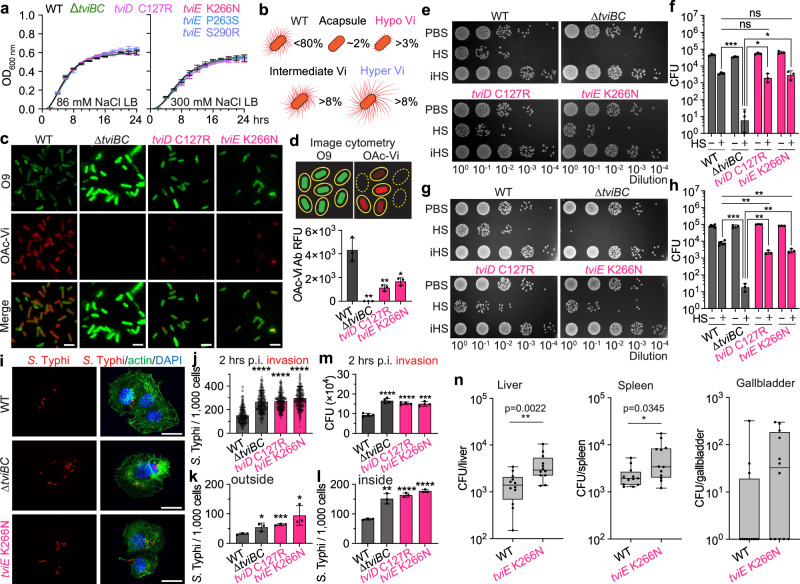
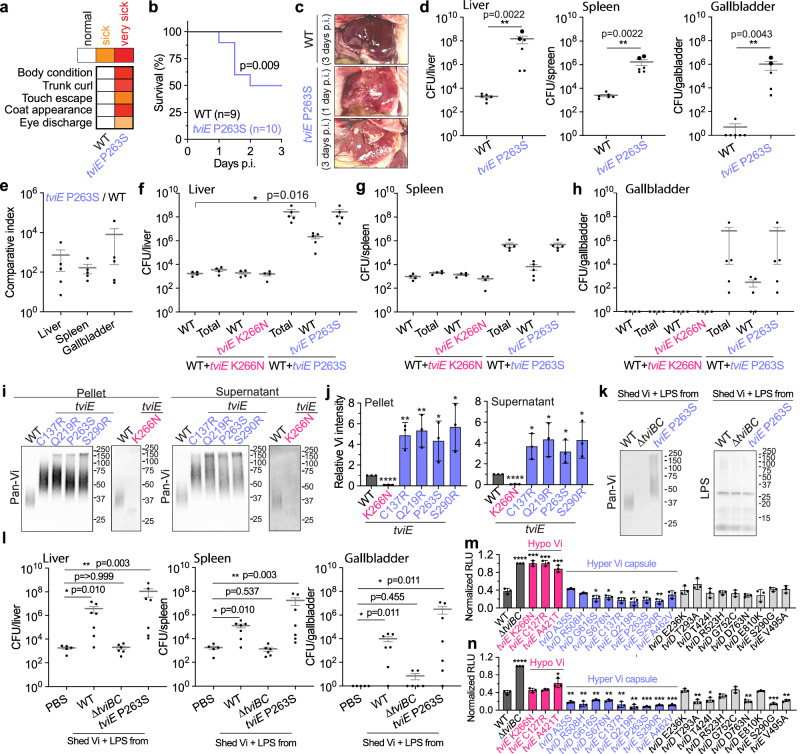
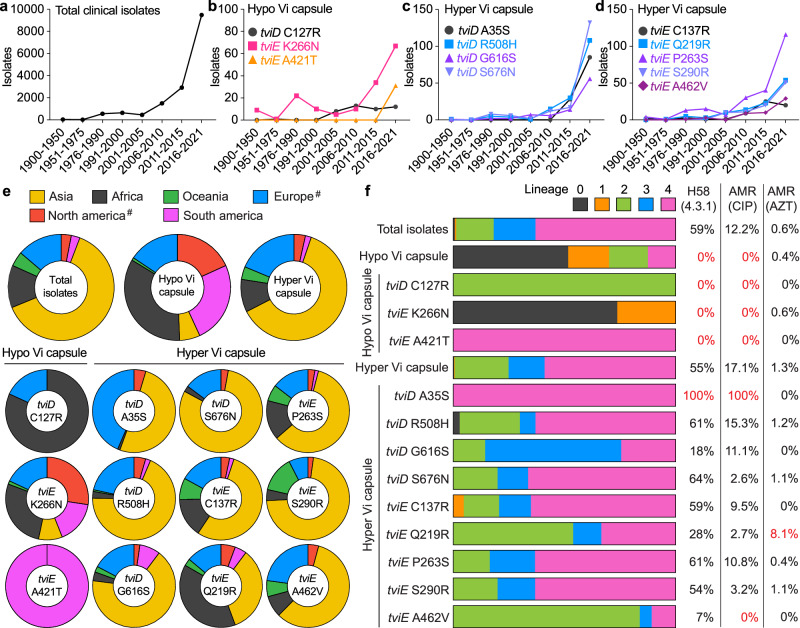
Many bacterial pathogens, including the human exclusive pathogen Salmonella Typhi, express capsular polysaccharides as a crucial virulence factor. Here, through S. Typhi whole genome sequence analyses and functional studies, we found a list of single point mutations that make S. Typhi hypervirulent. We discovered a single point mutation in the Vi biosynthesis enzymes that control Vi polymerization or acetylation is enough to result in different capsule variants of S. Typhi. All variant strains are pathogenic, but the hyper Vi capsule variants are particularly hypervirulent, as demonstrated by the high morbidity and mortality rates observed in infected mice. The hypo Vi capsule variants have primarily been identified in Africa, whereas the hyper Vi capsule variants are distributed worldwide. Collectively, these studies increase awareness about the existence of different capsule variants of S. Typhi, establish a solid foundation for numerous future studies on S. Typhi capsule variants, and offer valuable insights into strategies to combat capsulated bacteria.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Single missense mutations in Vi capsule synthesis genes confer hypervirulence to Salmonella Typhi.bioRxiv [Preprint]. 2024 Jan 3:2023.12.28.573590. doi: 10.1101/2023.12.28.573590. bioRxiv. 2024. Update in: Nat Commun. 2024 Jun 19;15(1):5258. doi: 10.1038/s41467-024-49590-6. PMID: 38260632 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- R01 AI139625/AI/NIAID NIH HHS/United States
- R01 AI141514/AI/NIAID NIH HHS/United States
- AI139625/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- AI141514/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
