

AMPK targets PDZD8 to trigger carbon source shift from glucose to glutamine
- PMID: 38898113
- PMCID: PMC11442470
- DOI: 10.1038/s41422-024-00985-6
AMPK targets PDZD8 to trigger carbon source shift from glucose to glutamine
Abstract
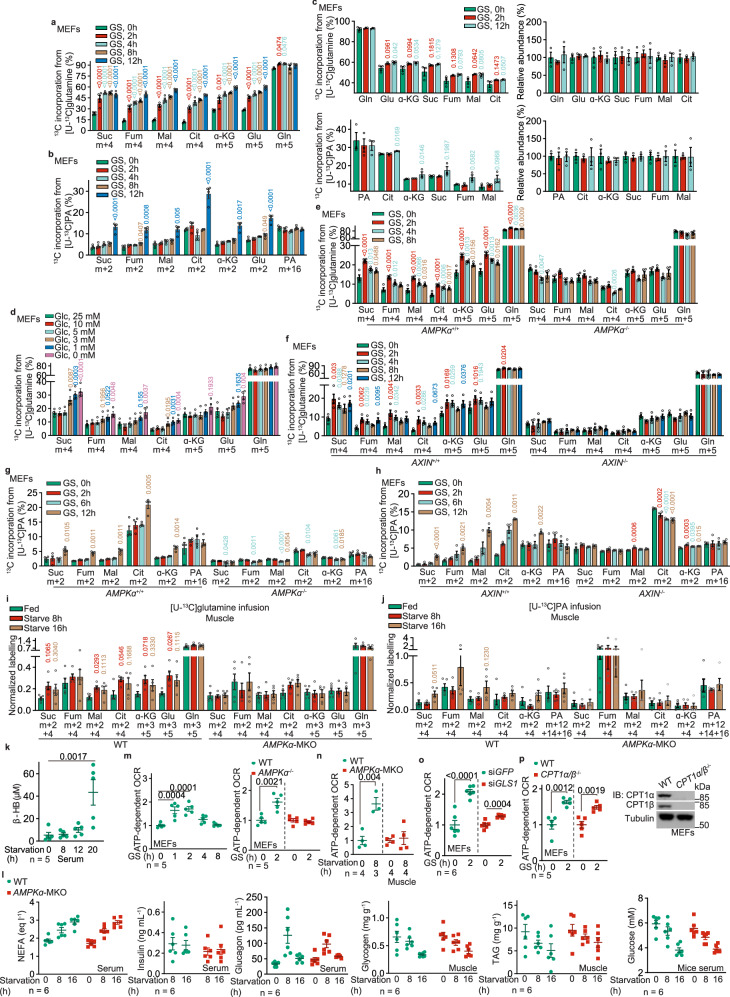
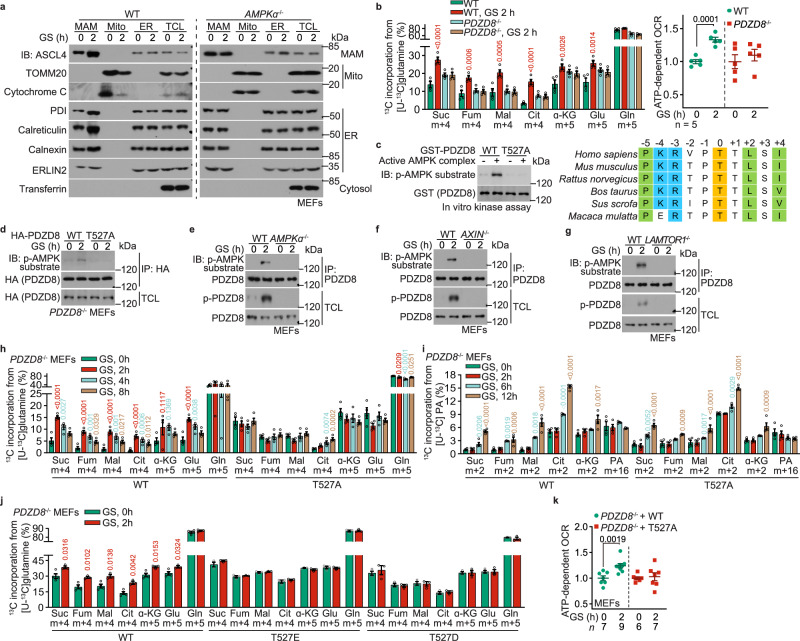
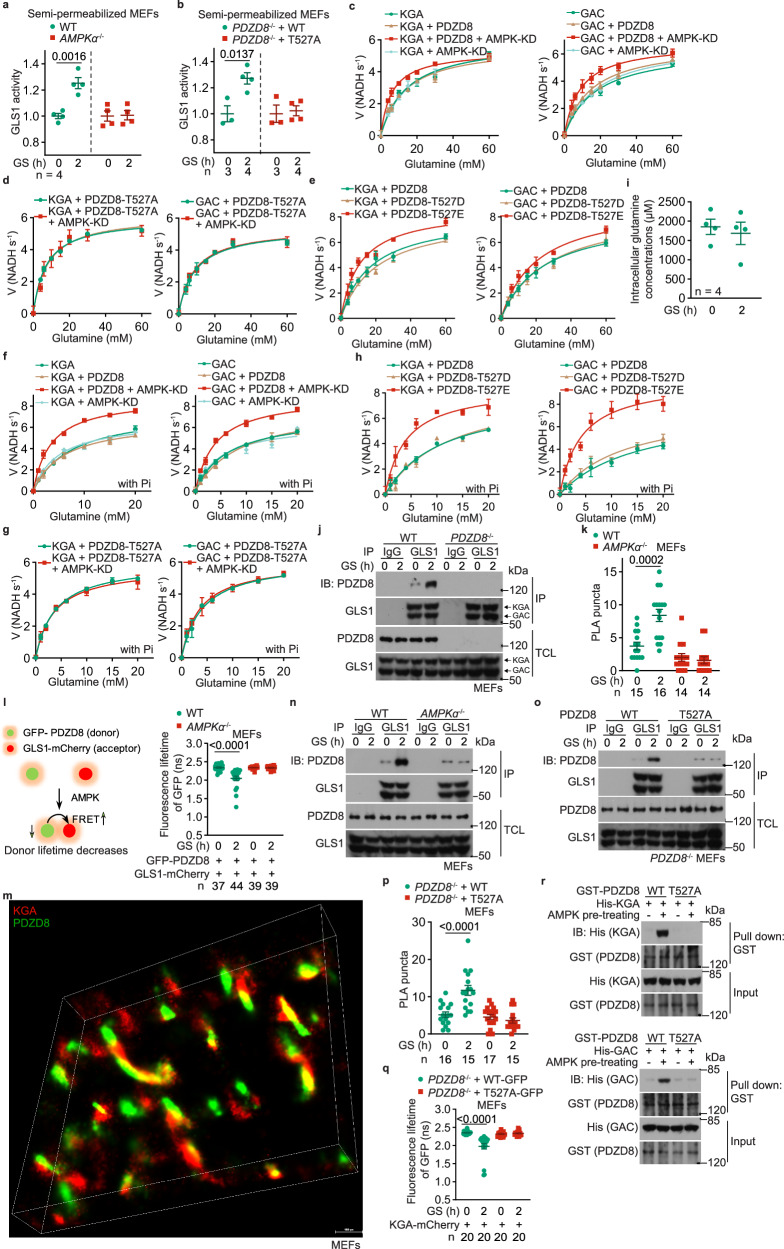
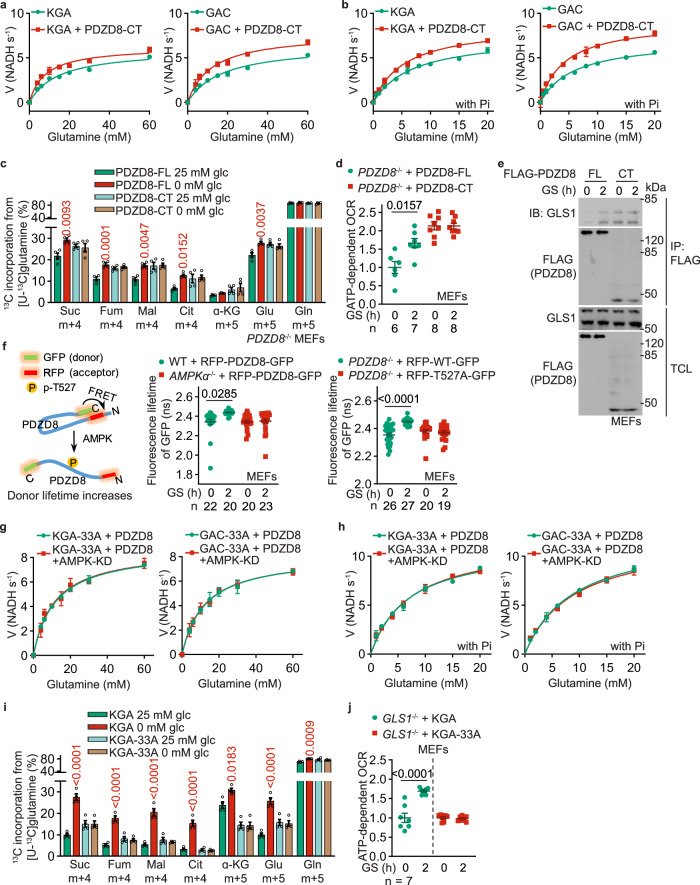
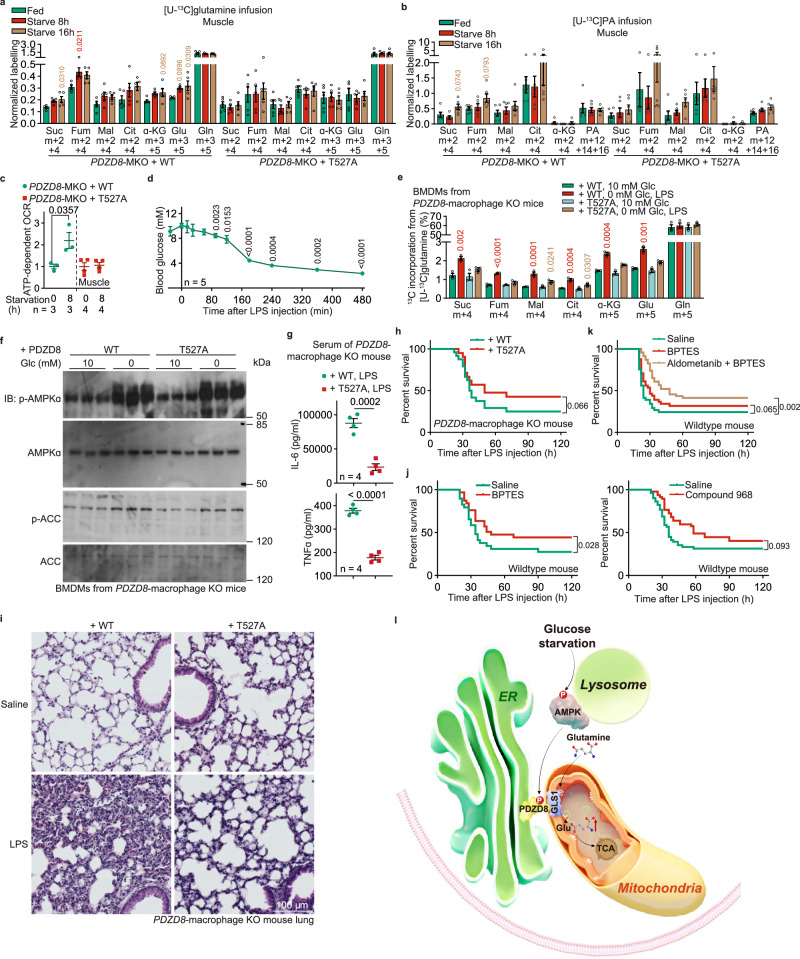
The shift of carbon utilization from primarily glucose to other nutrients is a fundamental metabolic adaptation to cope with decreased blood glucose levels and the consequent decline in glucose oxidation. AMP-activated protein kinase (AMPK) plays crucial roles in this metabolic adaptation. However, the underlying mechanism is not fully understood. Here, we show that PDZ domain containing 8 (PDZD8), which we identify as a new substrate of AMPK activated in low glucose, is required for the low glucose-promoted glutaminolysis. AMPK phosphorylates PDZD8 at threonine 527 (T527) and promotes the interaction of PDZD8 with and activation of glutaminase 1 (GLS1), a rate-limiting enzyme of glutaminolysis. In vivo, the AMPK-PDZD8-GLS1 axis is required for the enhancement of glutaminolysis as tested in the skeletal muscle tissues, which occurs earlier than the increase in fatty acid utilization during fasting. The enhanced glutaminolysis is also observed in macrophages in low glucose or under acute lipopolysaccharide (LPS) treatment. Consistent with a requirement of heightened glutaminolysis, the PDZD8-T527A mutation dampens the secretion of pro-inflammatory cytokines in macrophages in mice treated with LPS. Together, we have revealed an AMPK-PDZD8-GLS1 axis that promotes glutaminolysis ahead of increased fatty acid utilization under glucose shortage.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
- 82088102, 91854208/National Natural Science Foundation of China (National Science Foundation of China)
- 32070753/National Natural Science Foundation of China (National Science Foundation of China)
- 323B2035/National Natural Science Foundation of China (National Science Foundation of China)
- 31900542/National Natural Science Foundation of China (National Science Foundation of China)
- 20720200069/Xiamen University (XMU)
- 2021X1183, 2022Y1233/Xiamen University (XMU)
- BP2018017/Ministry of Education of the People's Republic of China (Ministry of Education of China)
- 2021Y9232/Natural Science Foundation of Fujian Province (Fujian Provincial Natural Science Foundation)
- 2023Y9448/Natural Science Foundation of Fujian Province (Fujian Provincial Natural Science Foundation)
- 2021Y9227/Natural Science Foundation of Fujian Province (Fujian Provincial Natural Science Foundation)
- 2020YFA0803402/Ministry of Science and Technology of the People's Republic of China (Chinese Ministry of Science and Technology)
- 2022ZD01005, 2022ZQNZD009, 2023L3020/Fujian Provincial Department of Science and Technology (Department of Science and Technology of Fujian Province)
- 4769/Agilent Technologies
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
