

The vertebrate retina: a window into the evolution of computation in the brain
- PMID: 38899158
- PMCID: PMC11183302
- DOI: 10.1016/j.cobeha.2024.101391
The vertebrate retina: a window into the evolution of computation in the brain
Abstract
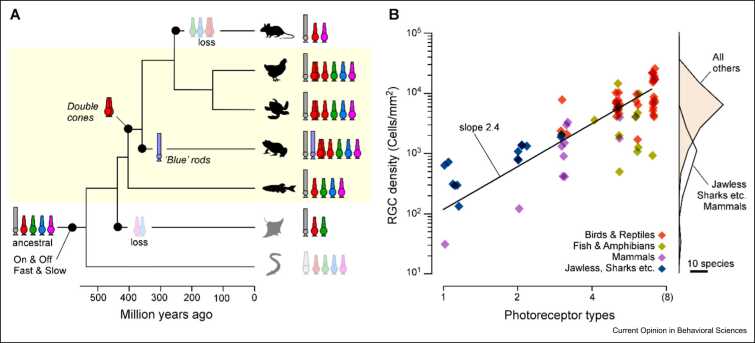
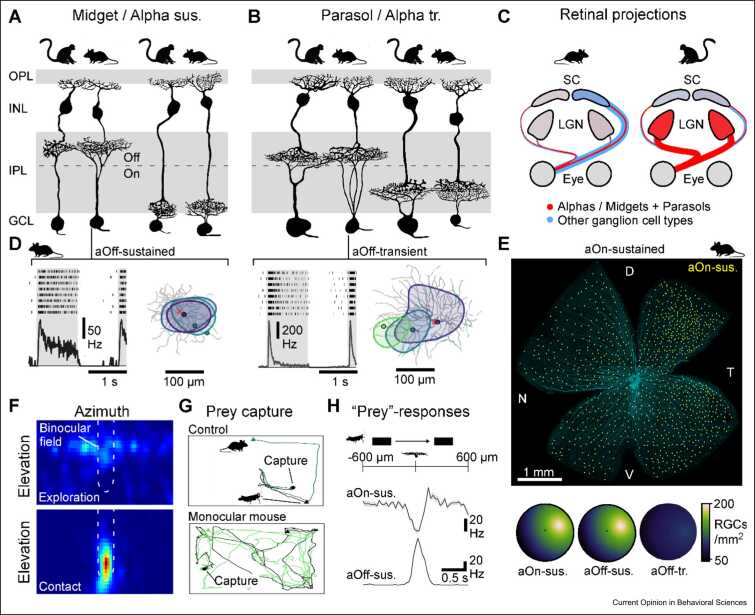
Animal brains are probably the most complex computational machines on our planet, and like everything in biology, they are the product of evolution. Advances in developmental and palaeobiology have been expanding our general understanding of how nervous systems can change at a molecular and structural level. However, how these changes translate into altered function - that is, into 'computation' - remains comparatively sparsely explored. What, concretely, does it mean for neuronal computation when neurons change their morphology and connectivity, when new neurons appear or old ones disappear, or when transmitter systems are slowly modified over many generations? And how does evolution use these many possible knobs and dials to constantly tune computation to give rise to the amazing diversity in animal behaviours we see today? Addressing these major gaps of understanding benefits from choosing a suitable model system. Here, I present the vertebrate retina as one perhaps unusually promising candidate. The retina is ancient and displays highly conserved core organisational principles across the entire vertebrate lineage, alongside a myriad of adjustments across extant species that were shaped by the history of their visual ecology. Moreover, the computational logic of the retina is readily interrogated experimentally, and our existing understanding of retinal circuits in a handful of species can serve as an anchor when exploring the visual circuit adaptations across the entire vertebrate tree of life, from fish deep in the aphotic zone of the oceans to eagles soaring high up in the sky.
© 2024 The Author(s).
Conflict of interest statement
None.
Figures


Similar articles
-
Energy limitation as a selective pressure on the evolution of sensory systems.J Exp Biol. 2008 Jun;211(Pt 11):1792-804. doi: 10.1242/jeb.017574. J Exp Biol. 2008. PMID: 18490395 Review.
-
A cell atlas of the chick retina based on single-cell transcriptomics.Elife. 2021 Jan 4;10:e63907. doi: 10.7554/eLife.63907. Elife. 2021. PMID: 33393903 Free PMC article.
-
Nonvisual photoreceptors of the deep brain, pineal organs and retina.Histol Histopathol. 2002 Apr;17(2):555-90. doi: 10.14670/HH-17.555. Histol Histopathol. 2002. PMID: 11962759 Review.
-
Development of the vertebrate retinal direction-selective circuit.Dev Biol. 2021 Sep;477:273-283. doi: 10.1016/j.ydbio.2021.06.004. Epub 2021 Jun 10. Dev Biol. 2021. PMID: 34118273 Free PMC article. Review.
-
Strip1 regulates retinal ganglion cell survival by suppressing Jun-mediated apoptosis to promote retinal neural circuit formation.Elife. 2022 Mar 22;11:e74650. doi: 10.7554/eLife.74650. Elife. 2022. PMID: 35314028 Free PMC article.
Cited by
-
Morphology and connectivity of retinal horizontal cells in two avian species.Front Cell Neurosci. 2025 Mar 4;19:1558605. doi: 10.3389/fncel.2025.1558605. eCollection 2025. Front Cell Neurosci. 2025. PMID: 40103750 Free PMC article.
-
Differential retinal ganglion cell resilience to optic nerve injury across vertebrate species.Front Neurosci. 2025 May 20;19:1596464. doi: 10.3389/fnins.2025.1596464. eCollection 2025. Front Neurosci. 2025. PMID: 40463588 Free PMC article.
-
Robust Sensory Traits Across Light Habitats: Visual Signals but Not Receptors Vary in Centrarchids Inhabiting Distinct Photic Environments.Mol Ecol. 2025 Apr;34(8):e17721. doi: 10.1111/mec.17721. Epub 2025 Mar 11. Mol Ecol. 2025. PMID: 40066691 Free PMC article.
References
-
- Roberts R.J.V., Pop S., Prieto-Godino L.L. Evolution of central neural circuits: state of the art and perspectives. Nat Rev Neurosci. 2022;23:725–743. - PubMed
-
State of the art summary of our understanding of the mechanisms by which neural circuits evolve.
-
- Kawecki T.J., et al. Experimental evolution. Trends Ecol Evol. 2012;27:547–560. - PubMed
-
- Baden T., Euler T., Berens P. Understanding the retinal basis of vision across species. Nat Rev Neurosci. 2020;21:5–20. - PubMed
-
- Baden T. Vertebrate vision: lessons from non-model species. Semin Cell Dev Biol. 2020;106:1–4. - PubMed
Publication types
LinkOut - more resources
Full Text Sources