

DYRK1A signalling synchronizes the mitochondrial import pathways for metabolic rewiring
- PMID: 38902238
- PMCID: PMC11189921
- DOI: 10.1038/s41467-024-49611-4
DYRK1A signalling synchronizes the mitochondrial import pathways for metabolic rewiring
Abstract
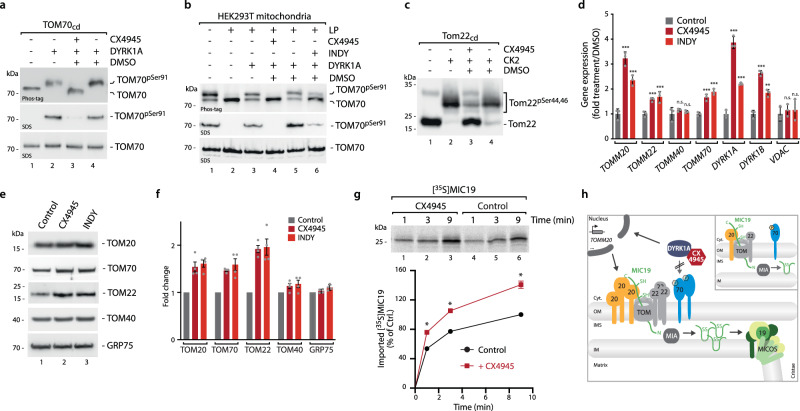
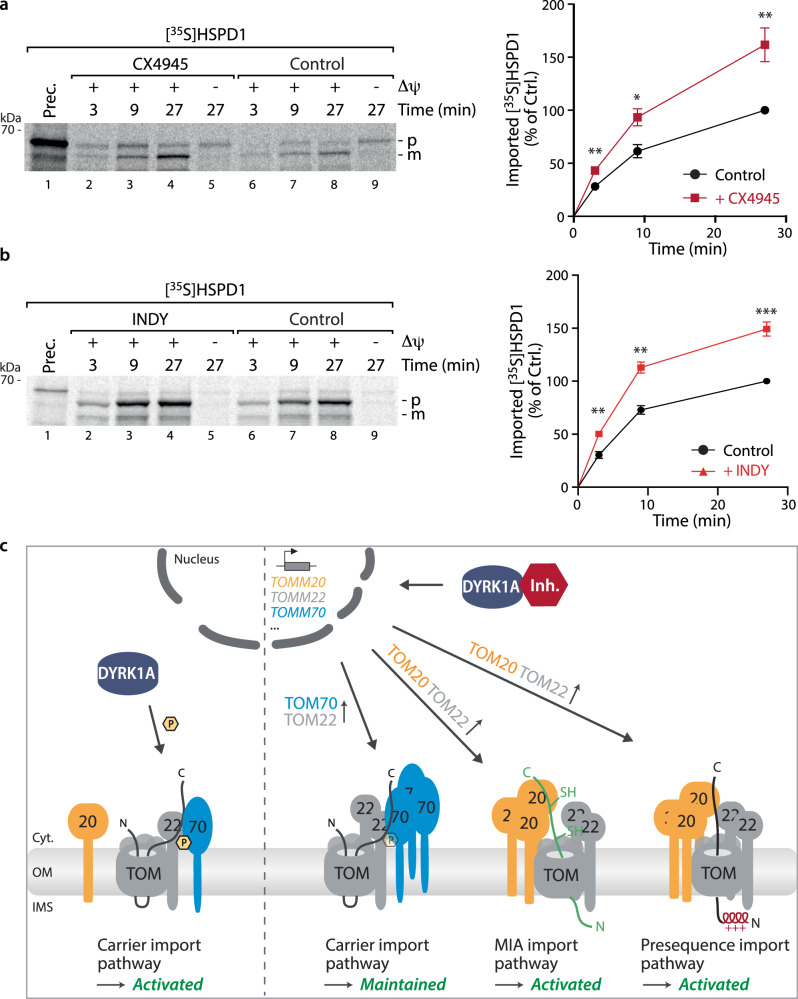
Mitochondria require an extensive proteome to maintain a variety of metabolic reactions, and changes in cellular demand depend on rapid adaptation of the mitochondrial protein composition. The TOM complex, the organellar entry gate for mitochondrial precursors in the outer membrane, is a target for cytosolic kinases to modulate protein influx. DYRK1A phosphorylation of the carrier import receptor TOM70 at Ser91 enables its efficient docking and thus transfer of precursor proteins to the TOM complex. Here, we probe TOM70 phosphorylation in molecular detail and find that TOM70 is not a CK2 target nor import receptor for MIC19 as previously suggested. Instead, we identify TOM20 as a MIC19 import receptor and show off-target inhibition of the DYRK1A-TOM70 axis with the clinically used CK2 inhibitor CX4945 which activates TOM20-dependent import pathways. Taken together, modulation of DYRK1A signalling adapts the central mitochondrial protein entry gate via synchronization of TOM70- and TOM20-dependent import pathways for metabolic rewiring. Thus, DYRK1A emerges as a cytosolic surveillance kinase to regulate and fine-tune mitochondrial protein biogenesis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests
Figures
Similar articles
-
Global kinome profiling reveals DYRK1A as critical activator of the human mitochondrial import machinery.Nat Commun. 2021 Jul 13;12(1):4284. doi: 10.1038/s41467-021-24426-9. Nat Commun. 2021. PMID: 34257281 Free PMC article.
-
Interaction between the human mitochondrial import receptors Tom20 and Tom70 in vitro suggests a chaperone displacement mechanism.J Biol Chem. 2011 Sep 16;286(37):32208-19. doi: 10.1074/jbc.M111.280446. Epub 2011 Jul 19. J Biol Chem. 2011. PMID: 21771790 Free PMC article.
-
Roles of Tom70 in import of presequence-containing mitochondrial proteins.J Biol Chem. 2009 Nov 13;284(46):31635-46. doi: 10.1074/jbc.M109.041756. Epub 2009 Sep 18. J Biol Chem. 2009. PMID: 19767391 Free PMC article.
-
Role of the TOM Complex in Protein Import into Mitochondria: Structural Views.Annu Rev Biochem. 2022 Jun 21;91:679-703. doi: 10.1146/annurev-biochem-032620-104527. Epub 2022 Mar 14. Annu Rev Biochem. 2022. PMID: 35287471 Review.
-
Coupling of import and assembly pathways in mitochondrial protein biogenesis.Biol Chem. 2019 Dec 18;401(1):117-129. doi: 10.1515/hsz-2019-0310. Biol Chem. 2019. PMID: 31513529 Review.
Cited by
-
Mesenchymal stem cell-derived small extracellular vesicles reduced hepatic lipid accumulation in MASLD by suppressing mitochondrial fission.Stem Cell Res Ther. 2025 Mar 5;16(1):116. doi: 10.1186/s13287-025-04228-2. Stem Cell Res Ther. 2025. PMID: 40045380 Free PMC article.
-
The mitochondrial intermembrane space - a permanently proteostasis-challenged compartment.Biol Chem. 2025 May 27;406(5-7):263-294. doi: 10.1515/hsz-2025-0108. Print 2025 Aug 26. Biol Chem. 2025. PMID: 40435180 Review.
References
MeSH terms
Substances
Grants and funding
- 403222702 SFB1381/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 390939984 EXC2189/Deutsche Forschungsgemeinschaft (German Research Foundation)
- RTG2202/Deutsche Forschungsgemeinschaft (German Research Foundation)
- Heisenberg Programm BR 3662/5/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 390939984 EXC2189/Deutsche Forschungsgemeinschaft (German Research Foundation)
LinkOut - more resources
Full Text Sources
