

This is a preprint.
An egg sabotaging mechanism drives non-Mendelian transmission in mice
- PMID: 38903120
- PMCID: PMC11188085
- DOI: 10.1101/2024.02.22.581453
An egg sabotaging mechanism drives non-Mendelian transmission in mice
Update in
-
An egg-sabotaging mechanism drives non-Mendelian transmission in mice.Curr Biol. 2024 Sep 9;34(17):3845-3854.e4. doi: 10.1016/j.cub.2024.07.001. Epub 2024 Jul 26. Curr Biol. 2024. PMID: 39067449 Free PMC article.
Abstract
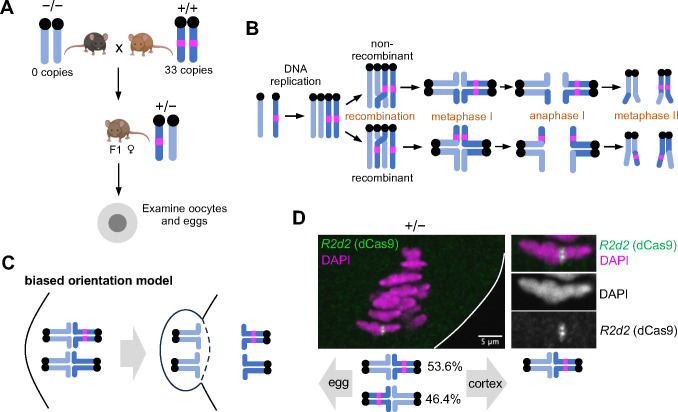
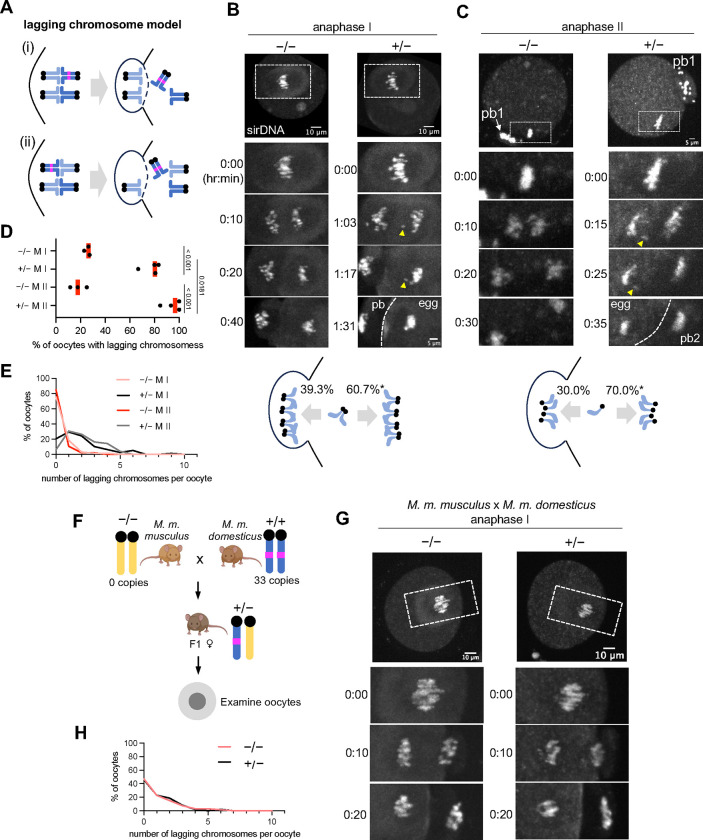
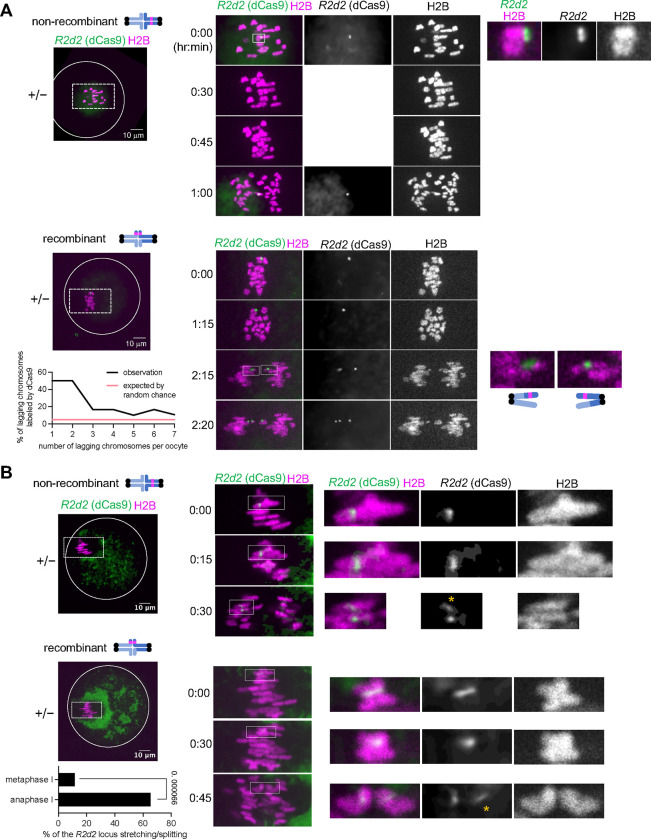
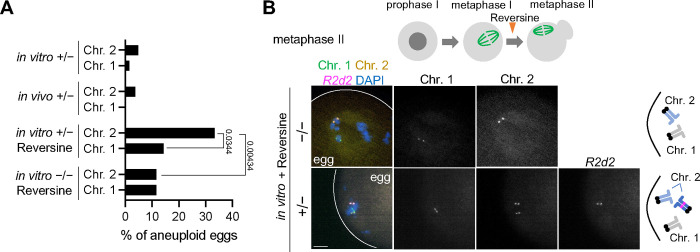
During meiosis, homologous chromosomes segregate so that alleles are transmitted equally to haploid gametes, following Mendel's Law of Segregation. However, some selfish genetic elements drive in meiosis to distort the transmission ratio and increase their representation in gametes. The established paradigms for drive are fundamentally different for female vs male meiosis. In male meiosis, selfish elements typically kill gametes that do not contain them. In female meiosis, killing is predetermined, and selfish elements bias their segregation to the single surviving gamete (i.e., the egg in animal meiosis). Here we show that a selfish element on mouse chromosome 2, R2d2, drives using a hybrid mechanism in female meiosis, incorporating elements of both male and female drivers. If R2d2 is destined for the polar body, it manipulates segregation to sabotage the egg by causing aneuploidy that is subsequently lethal in the embryo, so that surviving progeny preferentially contain R2d2. In heterozygous females, R2d2 orients randomly on the metaphase spindle but lags during anaphase and preferentially remains in the egg, regardless of its initial orientation. Thus, the egg genotype is either euploid with R2d2 or aneuploid with both homologs of chromosome 2, with only the former generating viable embryos. Consistent with this model, R2d2 heterozygous females produce eggs with increased aneuploidy for chromosome 2, increased embryonic lethality, and increased transmission of R2d2. In contrast to a male meiotic driver, which kills its sister gametes produced as daughter cells in the same meiosis, R2d2 eliminates "cousins" produced from meioses in which it should have been excluded from the egg.
Keywords: aneuploidy; chromosome segregation; female meiotic drive; meiosis; mouse oocyte; selfish genetic element.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources