

TERT activation targets DNA methylation and multiple aging hallmarks
- PMID: 38908367
- PMCID: PMC11552617
- DOI: 10.1016/j.cell.2024.05.048
TERT activation targets DNA methylation and multiple aging hallmarks
Abstract
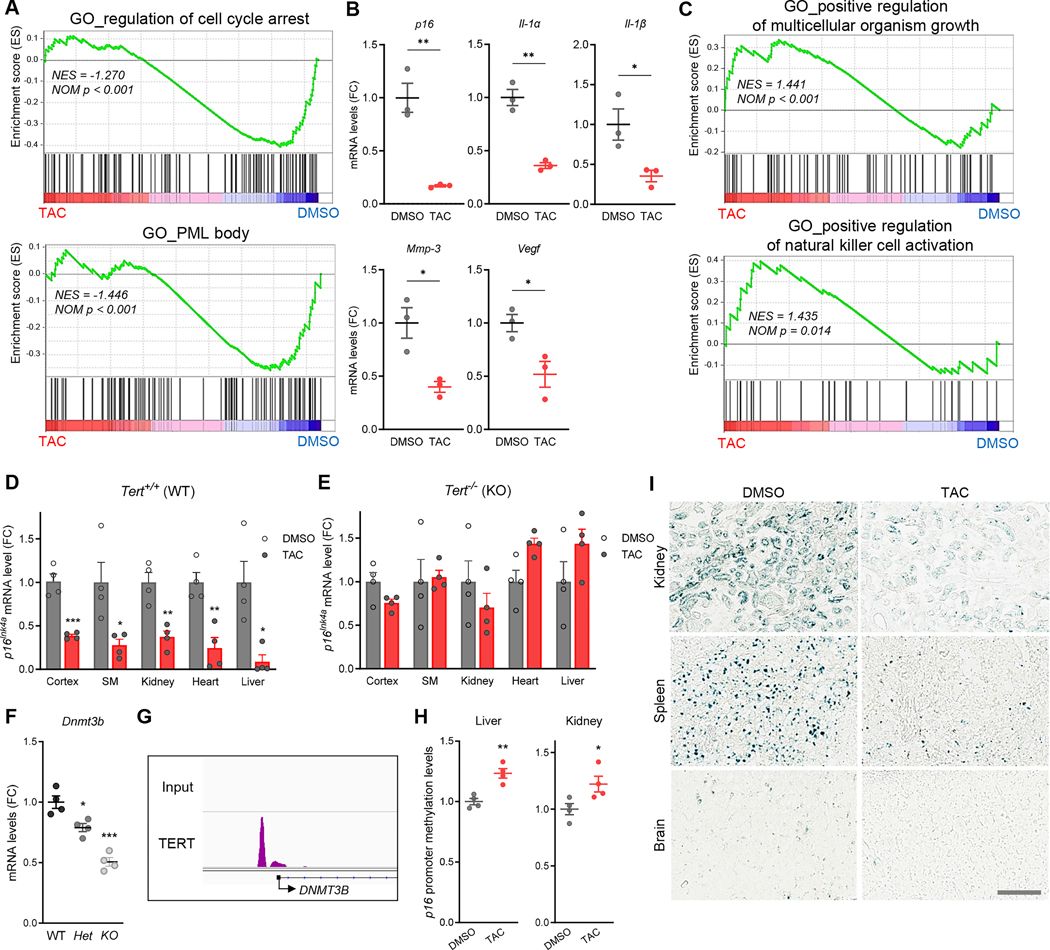
Insufficient telomerase activity, stemming from low telomerase reverse transcriptase (TERT) gene transcription, contributes to telomere dysfunction and aging pathologies. Besides its traditional function in telomere synthesis, TERT acts as a transcriptional co-regulator of genes pivotal in aging and age-associated diseases. Here, we report the identification of a TERT activator compound (TAC) that upregulates TERT transcription via the MEK/ERK/AP-1 cascade. In primary human cells and naturally aged mice, TAC-induced elevation of TERT levels promotes telomere synthesis, blunts tissue aging hallmarks with reduced cellular senescence and inflammatory cytokines, and silences p16INK4a expression via upregulation of DNMT3B-mediated promoter hypermethylation. In the brain, TAC alleviates neuroinflammation, increases neurotrophic factors, stimulates adult neurogenesis, and preserves cognitive function without evident toxicity, including cancer risk. Together, these findings underscore TERT's critical role in aging processes and provide preclinical proof of concept for physiological TERT activation as a strategy to mitigate multiple aging hallmarks and associated pathologies.
Keywords: adult neurogenesis; cognition; epigenetics; inflammation; p16(INK4a); senescence; telomerase; telomere.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.A.D. is a founder, advisor, and/or director of Tvardi Therapeutics, Inc.; Nirogy Therapeutics, Inc.; Stellanova Therapeutics, Inc.; Sporos Bioventures, LLC.; Bectas Therapeutics, Inc.; and Asylia Therapeutics, Inc., which are focused on therapies for cancer and fibrosis and bear no direct relevance to this study.
Figures




References
-
- Carmona JJ, and Michan S. (2016). Biology of Healthy Aging and Longevity. Rev Invest Clin 68, 7–16. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
