

Inverted triplications formed by iterative template switches generate structural variant diversity at genomic disorder loci
- PMID: 38908378
- PMCID: PMC11293582
- DOI: 10.1016/j.xgen.2024.100590
Inverted triplications formed by iterative template switches generate structural variant diversity at genomic disorder loci
Abstract
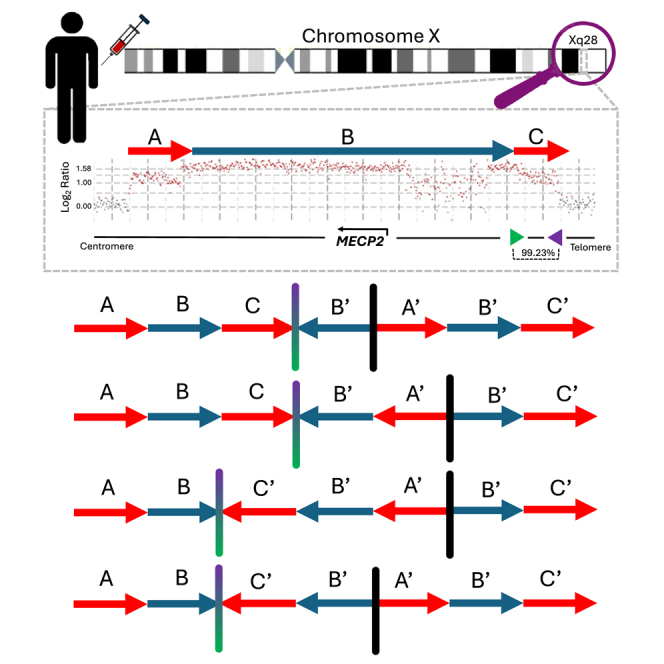
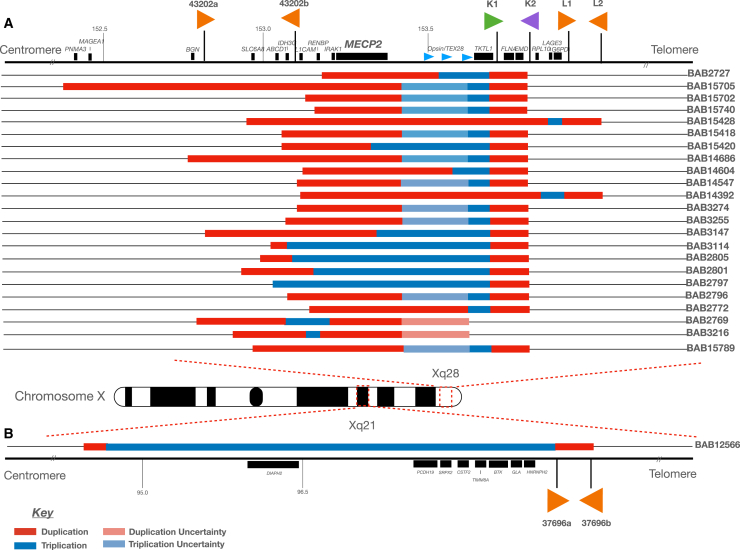
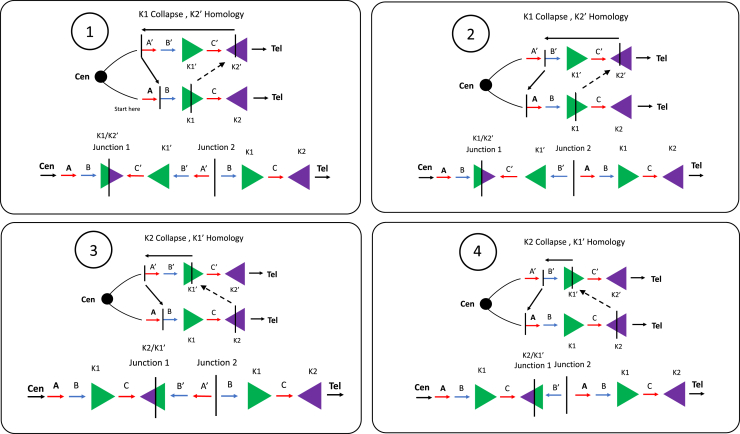
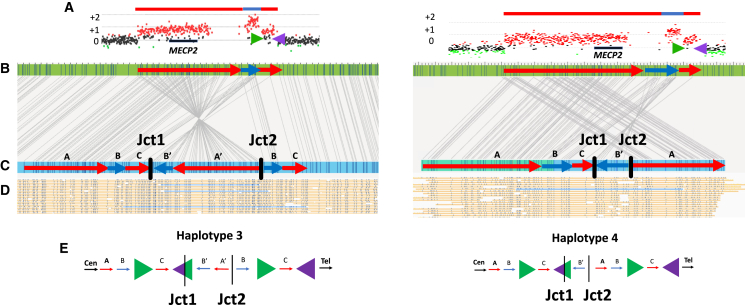
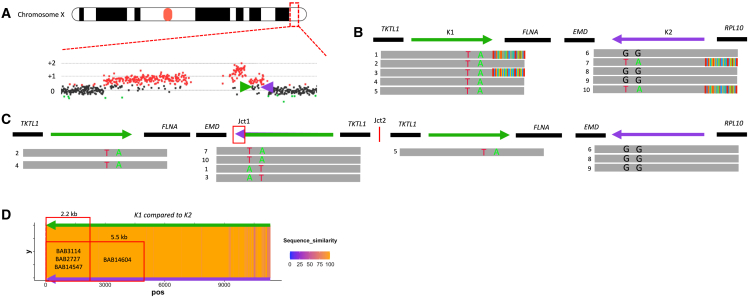
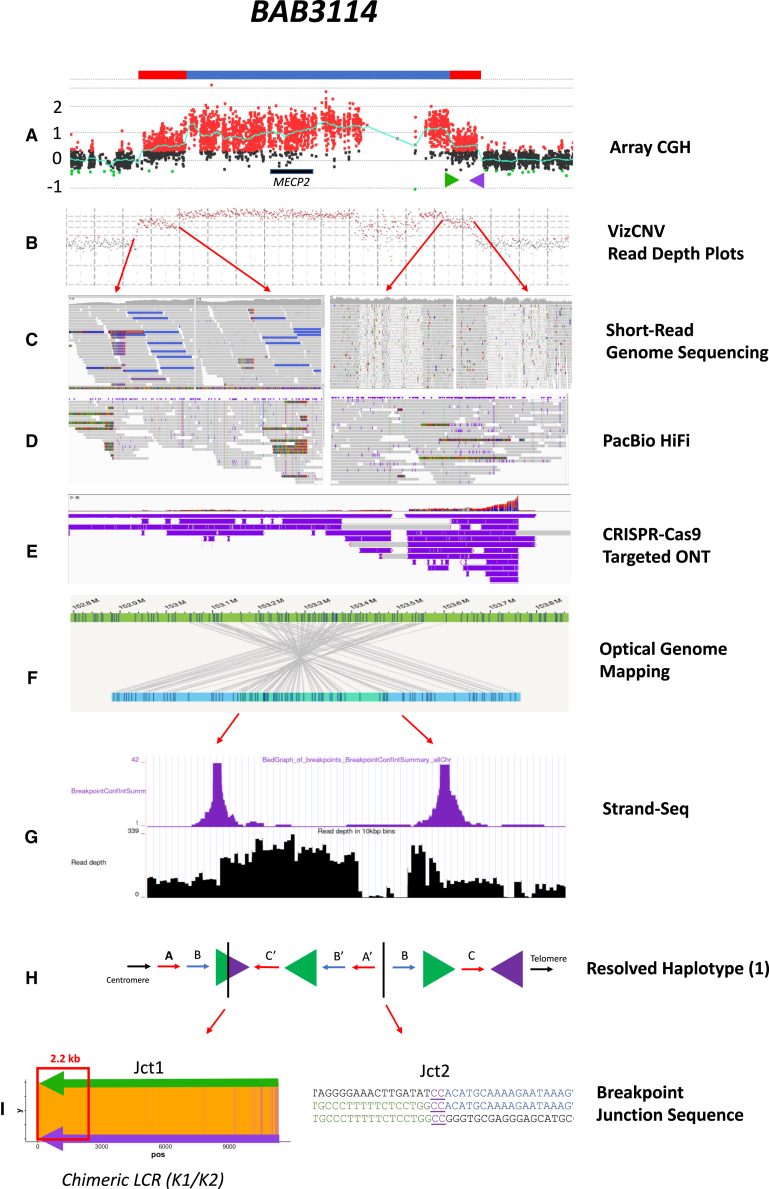
The duplication-triplication/inverted-duplication (DUP-TRP/INV-DUP) structure is a complex genomic rearrangement (CGR). Although it has been identified as an important pathogenic DNA mutation signature in genomic disorders and cancer genomes, its architecture remains unresolved. Here, we studied the genomic architecture of DUP-TRP/INV-DUP by investigating the DNA of 24 patients identified by array comparative genomic hybridization (aCGH) on whom we found evidence for the existence of 4 out of 4 predicted structural variant (SV) haplotypes. Using a combination of short-read genome sequencing (GS), long-read GS, optical genome mapping, and single-cell DNA template strand sequencing (strand-seq), the haplotype structure was resolved in 18 samples. The point of template switching in 4 samples was shown to be a segment of ∼2.2-5.5 kb of 100% nucleotide similarity within inverted repeat pairs. These data provide experimental evidence that inverted low-copy repeats act as recombinant substrates. This type of CGR can result in multiple conformers generating diverse SV haplotypes in susceptible dosage-sensitive loci.
Keywords: MECP2 duplication syndrome; MMBIR; Mendelian diseases; Xq28; break-induced replication; copy-number variant; inversions; recombination; segmental duplication; template switching.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Baylor College of Medicine and Miraca Holdings have formed a joint venture with shared ownership and governance of BG, which performs clinical microarray analysis, clinical ES, and clinical biochemical studies. J.R.L. serves on the scientific advisory board of the BG. J.R.L. has stock ownership in 23andMe, is a paid consultant for Genomics International, and is a co-inventor on multiple US and European patents related to molecular diagnostics for inherited neuropathies, eye diseases, genomic disorders, and bacterial genomic fingerprinting. E.H. and S.J. are employees of ONT and shareholders and/or share option holders of ONT. D.P. provides consulting services for Ionis Pharmaceuticals. F.J.S. receives research support from Genetech, Illumina, Pacbio, and ONT.
Figures
References
-
- Pellestor F., Anahory T., Lefort G., Puechberty J., Liehr T., Hédon B., Sarda P. Complex chromosomal rearrangements: origin and meiotic behavior. Hum. Reprod. Update. 2011;17:476–494. - PubMed
-
- Carvalho C.M.B., Ramocki M.B., Pehlivan D., Franco L.M., Gonzaga-Jauregui C., Fang P., McCall A., Pivnick E.K., Hines-Dowell S., Seaver L.H., et al. Inverted genomic segments and complex triplication rearrangements are mediated by inverted repeats in the human genome. Nat. Genet. 2011;43:1074–1081. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
