

Interactomic exploration of LRRC8A in volume-regulated anion channels
- PMID: 38909013
- PMCID: PMC11193767
- DOI: 10.1038/s41420-024-02032-0
Interactomic exploration of LRRC8A in volume-regulated anion channels
Erratum in
-
Publisher Correction: Interactomic exploration of LRRC8A in volume-regulated anion channels.Cell Death Discov. 2024 Aug 29;10(1):387. doi: 10.1038/s41420-024-02123-y. Cell Death Discov. 2024. PMID: 39209805 Free PMC article. No abstract available.
Abstract
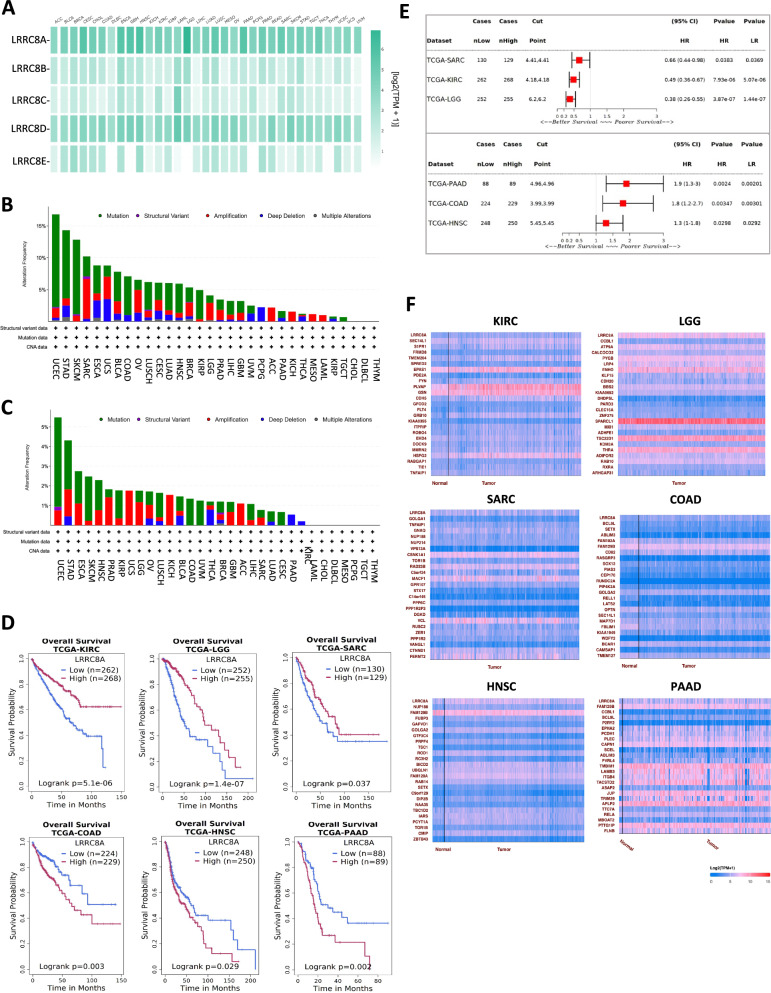
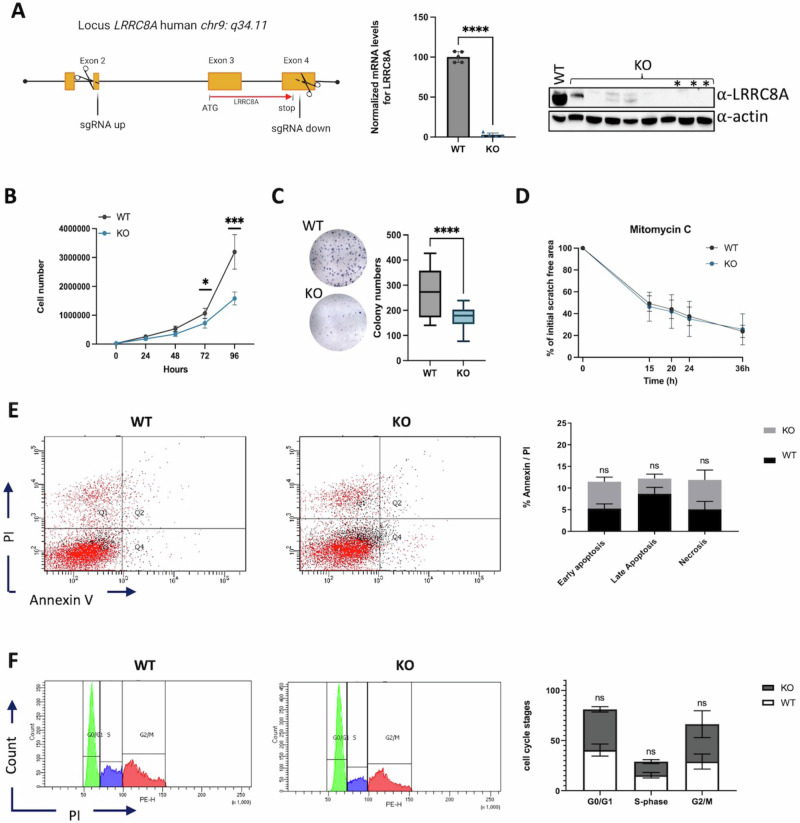
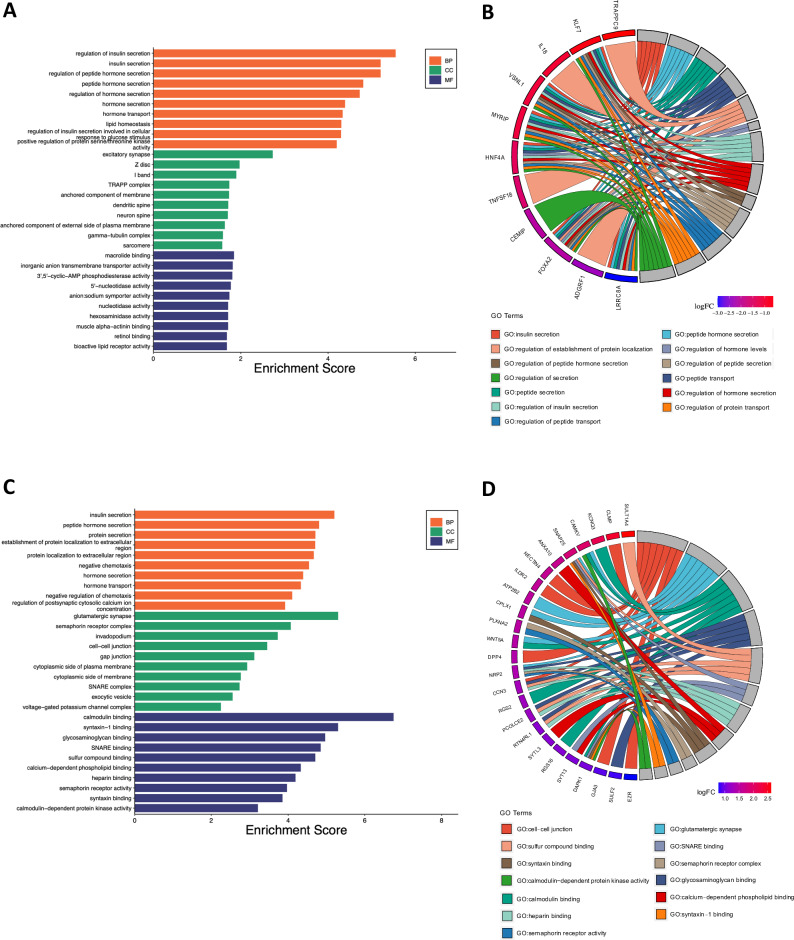
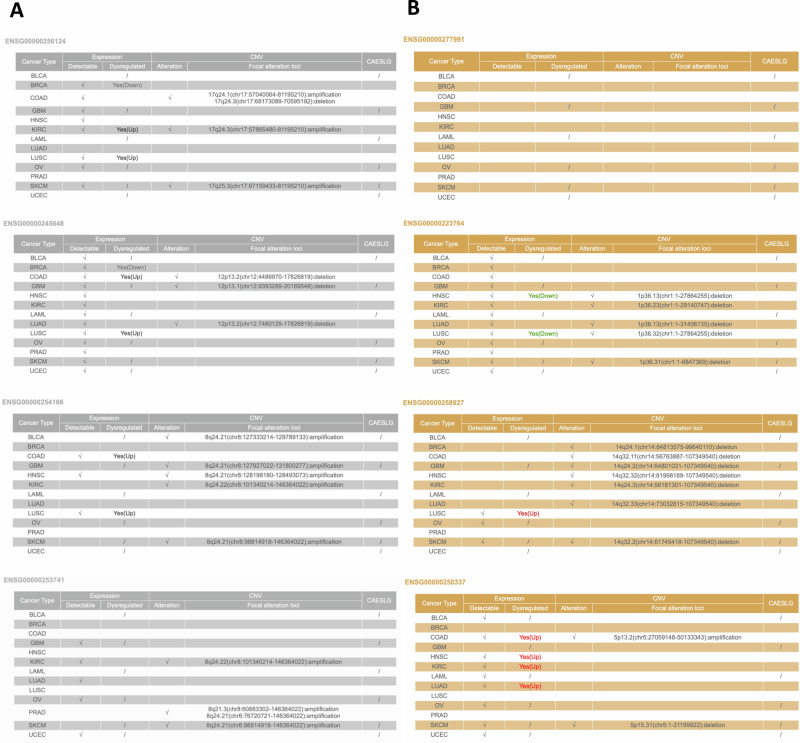
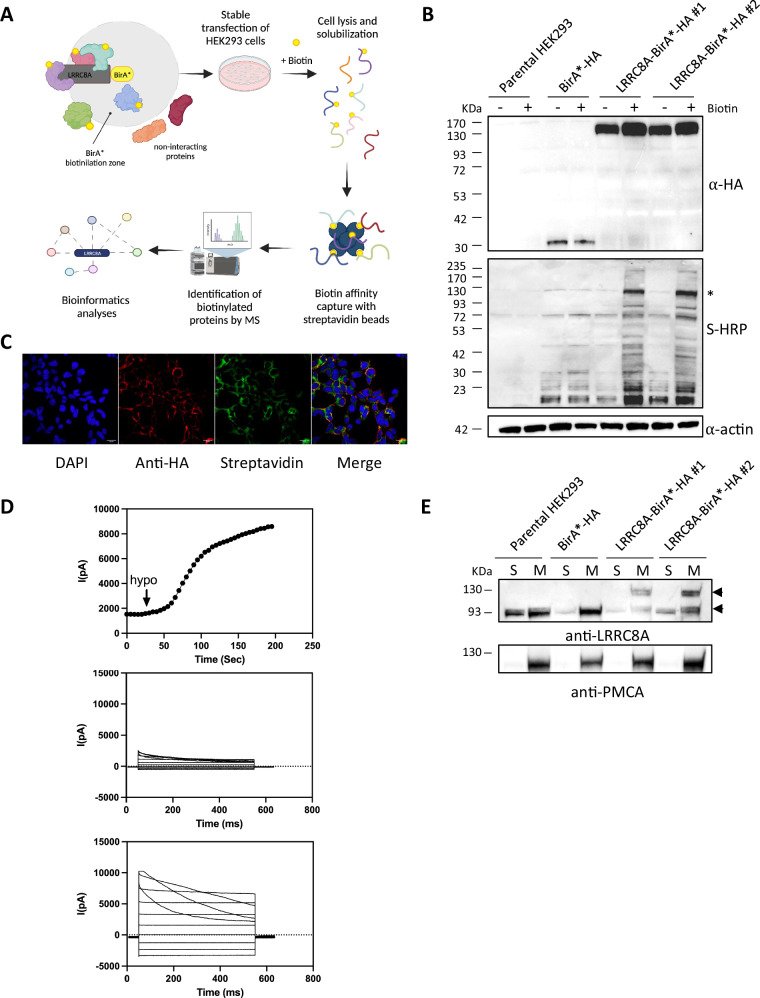
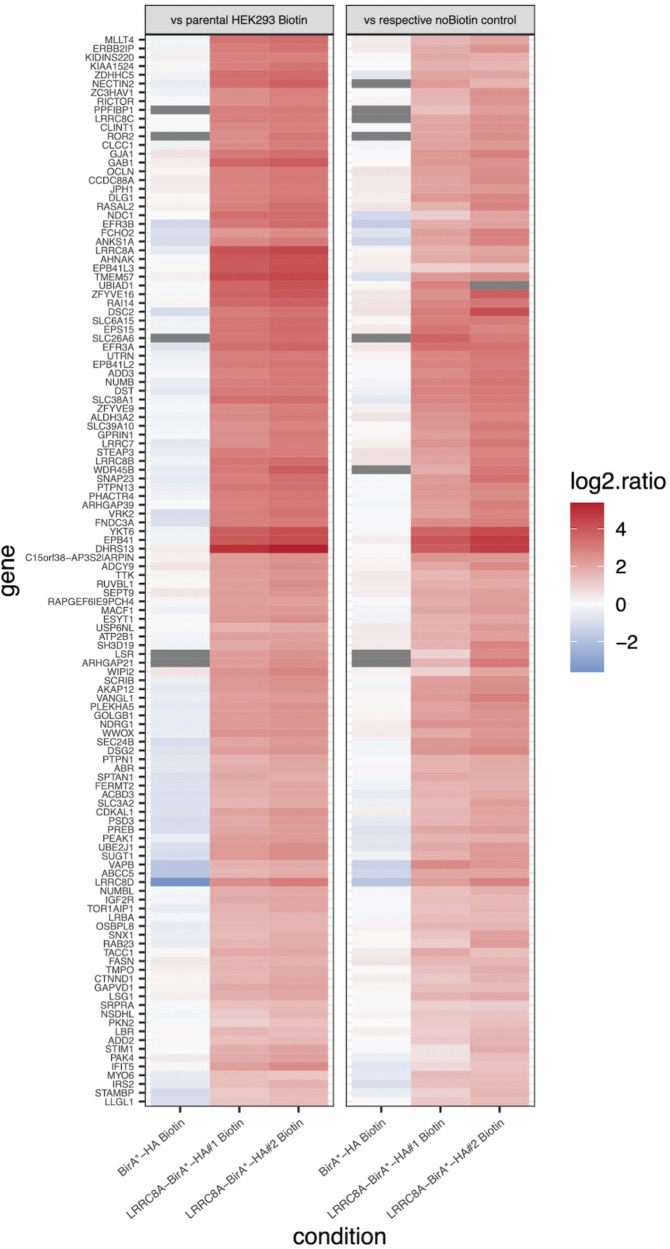
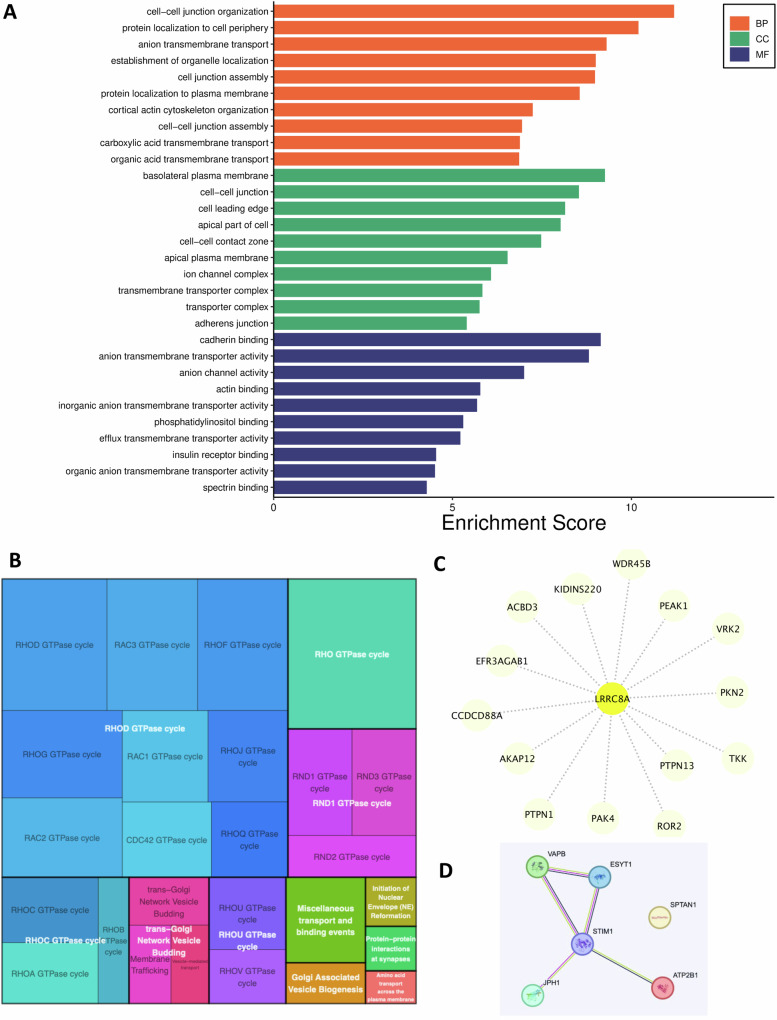
Ion channels are critical in enabling ion movement into and within cells and are important targets for pharmacological interventions in different human diseases. In addition to their ion transport abilities, ion channels interact with signalling and scaffolding proteins, which affects their function, cellular positioning, and links to intracellular signalling pathways. The study of "channelosomes" within cells has the potential to uncover their involvement in human diseases, although this field of research is still emerging. LRRC8A is the gene that encodes a crucial protein involved in the formation of volume-regulated anion channels (VRACs). Some studies suggest that LRRC8A could be a valuable prognostic tool in different types of cancer, serving as a biomarker for predicting patients' outcomes. LRRC8A expression levels might be linked to tumour progression, metastasis, and treatment response, although its implications in different cancer types can be varied. Here, publicly accessible databases of cancer patients were systematically analysed to determine if a correlation between VRAC channel expression and survival rate exists across distinct cancer types. Moreover, we re-evaluated the impact of LRRC8A on cellular proliferation and migration in colon cancer via HCT116 LRRC8A-KO cells, which is a current topic of debate in the literature. In addition, to investigate the role of LRRC8A in cellular signalling, we conducted biotin proximity-dependent identification (BioID) analysis, revealing a correlation between VRAC channels and cell-cell junctions, mechanisms that govern cellular calcium homeostasis, kinases, and GTPase signalling. Overall, this dataset improves our understanding of LRRC8A/VRAC and explores new research avenues while identifying promising therapeutic targets and promoting inventive methods for disease treatment.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Grants and funding
- DOR (Dotazione Ordinaria della Ricerca dipartimentale) projects - Calls 2021-2022-2023/Università degli Studi di Padova (University of Padova)
- SEED-PRID project 2021 entitled "Setting up in vivo interactome for cancer research"/Università degli Studi di Padova (University of Padova)
- BIRD-PRID, grant number BIRD239198/23/Università degli Studi di Padova (University of Padova)
- PRIN, grant number 2022ZY7ATN/Ministero dell'Istruzione, dell'Università e della Ricerca (Ministry of Education, University and Research)
LinkOut - more resources
Full Text Sources
Research Materials
