

T cell independent antibody responses with class switch and memory using peptides anchored on liposomes
- PMID: 38909055
- PMCID: PMC11193769
- DOI: 10.1038/s41541-024-00902-3
T cell independent antibody responses with class switch and memory using peptides anchored on liposomes
Abstract
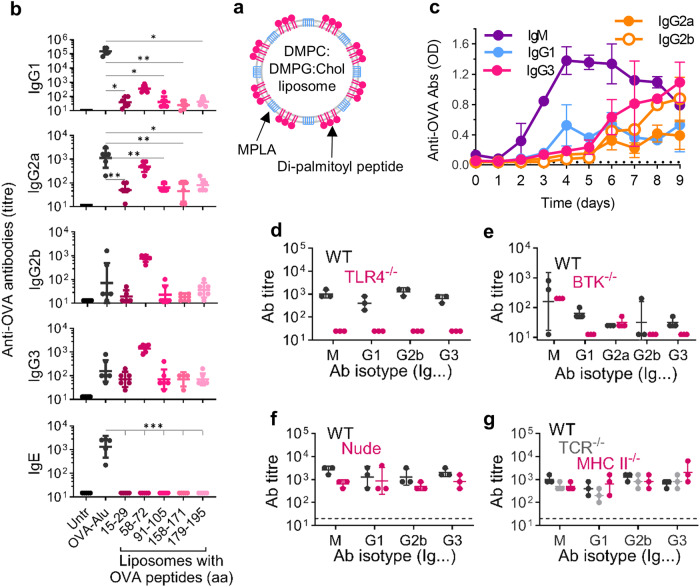
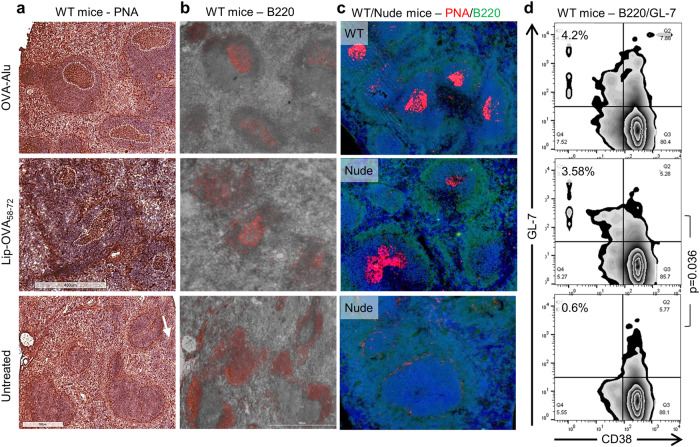
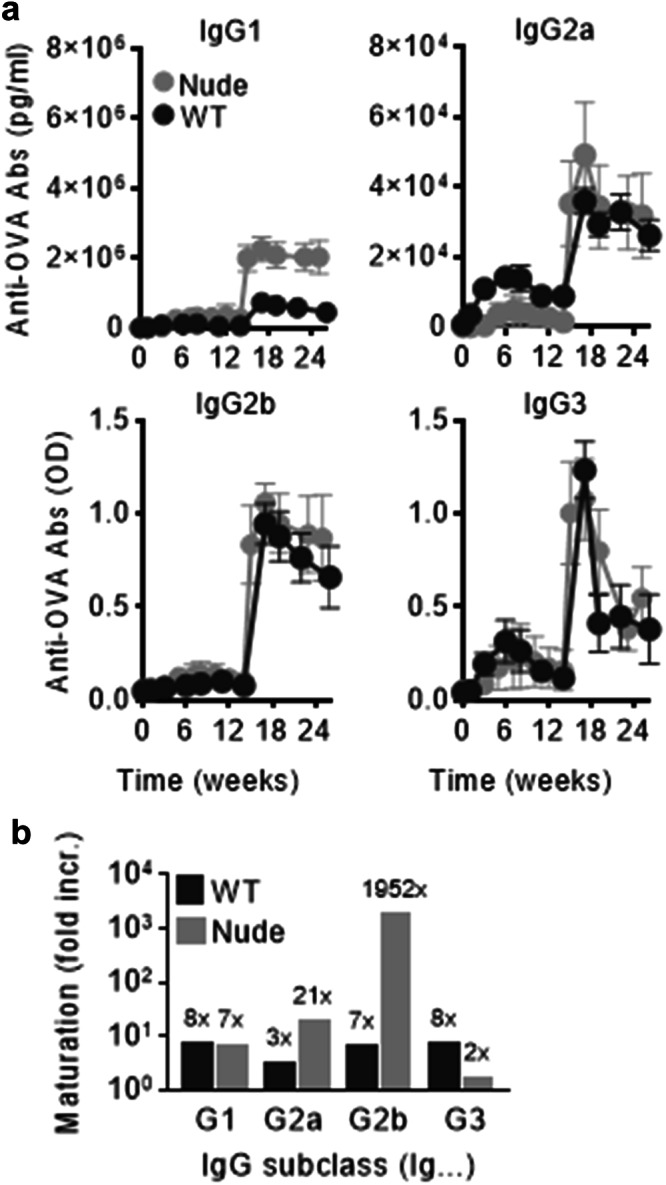
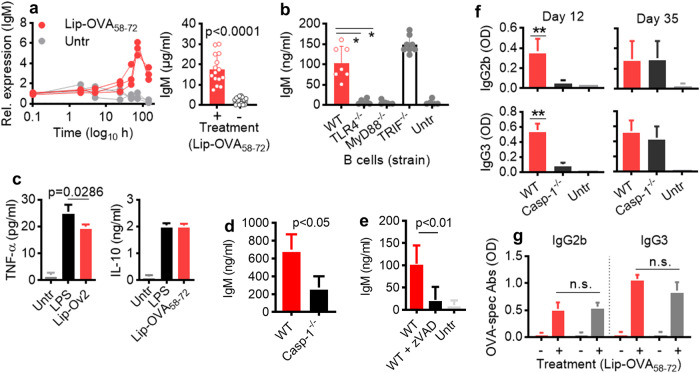
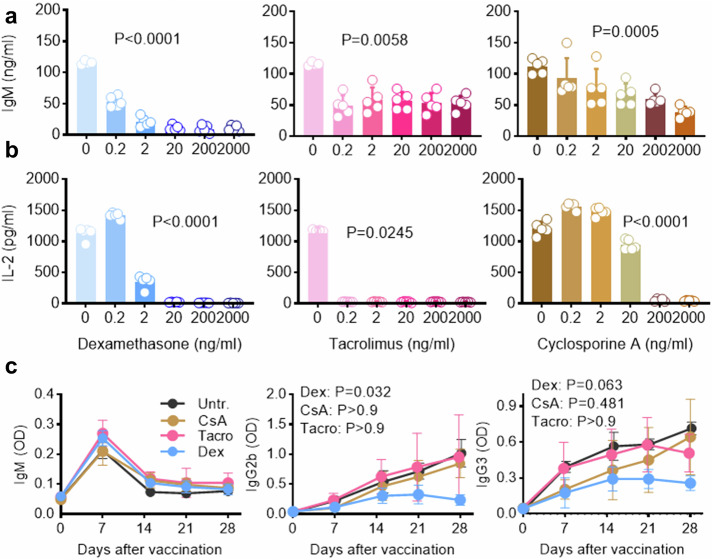
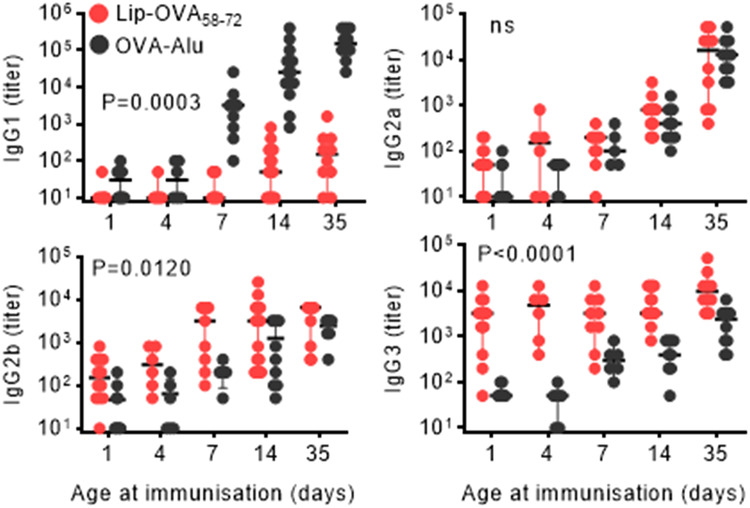
Vaccines generally require T lymphocytes for B-cell activation and immunoglobulin class switching in response to peptide or protein antigens. In the absence of T cells, limited IgG class switch takes place, germinal centers are short-lived, and the B cells lack memory. Here, immunization of mice with liposomes containing 15mer peptides and monophosphoryl lipid A (MPLA) as adjuvant, induced T-cell independent (TI) IgG class switch within three days, as well as germinal center formation. The antibody responses were long-lived, strictly dependent on Toll-like receptor 4 (TLR4) signaling, partly dependent on Bruton's tyrosine kinase (BTK) signal transmission, and independent of signaling through T-cell receptors, MHC class II and inflammasome. The antibody response showed characteristics of both TI type 1 and TI type 2. All IgG subclasses could be boosted months after primary immunization, and the biological function of the secreted antibodies was demonstrated in murine models of allergic anaphylaxis and of bacterial infection. Moreover, antibody responses after immunization with peptide- and MPLA-loaded liposomes could be triggered in neonatal mice and in mice receiving immune-suppressants. This study demonstrates T-cell independent endogenous B-cell memory and recall responses in vivo using a peptide antigen. The stimulation of these antibody responses required a correct and dense assembly and administration of peptide and adjuvant on the surface of liposomes. In the future, TI vaccines may prove beneficial in pathological conditions in which T-cell immunity is compromised through disease or medicines or when rapid, antibody-mediated immune protection is needed.
© 2024. The Author(s).
Conflict of interest statement
M.V. and M.P.L.D. are employed by AC Immune. M.P. is former employee of AC Immune. T.M.K. was scientific advisor to AC Immune. P.J. received material support from AC Immune. The authors declare that there are no further competing interests.
Figures


References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
