

Prefrontal coding of learned and inferred knowledge during REM and NREM sleep
- PMID: 38914541
- PMCID: PMC11196720
- DOI: 10.1038/s41467-024-48816-x
Prefrontal coding of learned and inferred knowledge during REM and NREM sleep
Abstract
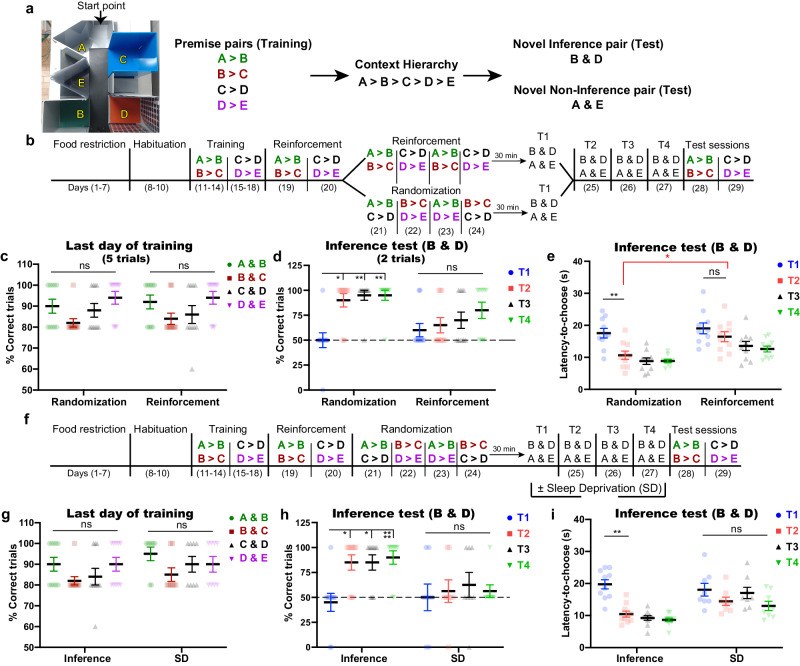
Idling brain activity has been proposed to facilitate inference, insight, and innovative problem-solving. However, it remains unclear how and when the idling brain can create novel ideas. Here, we show that cortical offline activity is both necessary and sufficient for building unlearned inferential knowledge from previously acquired information. In a transitive inference paradigm, male C57BL/6J mice gained the inference 1 day after, but not shortly after, complete training. Inhibiting the neuronal computations in the anterior cingulate cortex (ACC) during post-learning either non-rapid eye movement (NREM) or rapid eye movement (REM) sleep, but not wakefulness, disrupted the inference without affecting the learned knowledge. In vivo Ca2+ imaging suggests that NREM sleep organizes the scattered learned knowledge in a complete hierarchy, while REM sleep computes the inferential information from the organized hierarchy. Furthermore, after insufficient learning, artificial activation of medial entorhinal cortex-ACC dialog during only REM sleep created inferential knowledge. Collectively, our study provides a mechanistic insight on NREM and REM coordination in weaving inferential knowledge, thus highlighting the power of idling brain in cognitive flexibility.
© 2024. The Author(s).
Conflict of interest statement
S.M. owns equity in a company, Gene Therapy Research Institution, that commercializes the use of AAV vectors for gene therapy applications. To the extent that the work in this manuscript increases the value of these commercial holdings, S.M. has a conflict of interest. The other authors declare no competing interests.
Figures




References
MeSH terms
Grants and funding
- JP18H05213/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP23H05476/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP19K16892/MEXT | Japan Society for the Promotion of Science (JSPS)
- 20H03554/MEXT | Japan Society for the Promotion of Science (JSPS)
- 23H02785/MEXT | Japan Society for the Promotion of Science (JSPS)
- JPMJCR13W1/MEXT | JST | Core Research for Evolutional Science and Technology (CREST)
- JPMJCR23N2/MEXT | JST | Core Research for Evolutional Science and Technology (CREST)
- JP25115002/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- JP23gm6510028/Japan Agency for Medical Research and Development (AMED)
LinkOut - more resources
Full Text Sources
Miscellaneous
