

This is a preprint.
Visual neurons recognize complex image transformations
- PMID: 38915552
- PMCID: PMC11195111
- DOI: 10.1101/2024.06.10.598314
Visual neurons recognize complex image transformations
Abstract
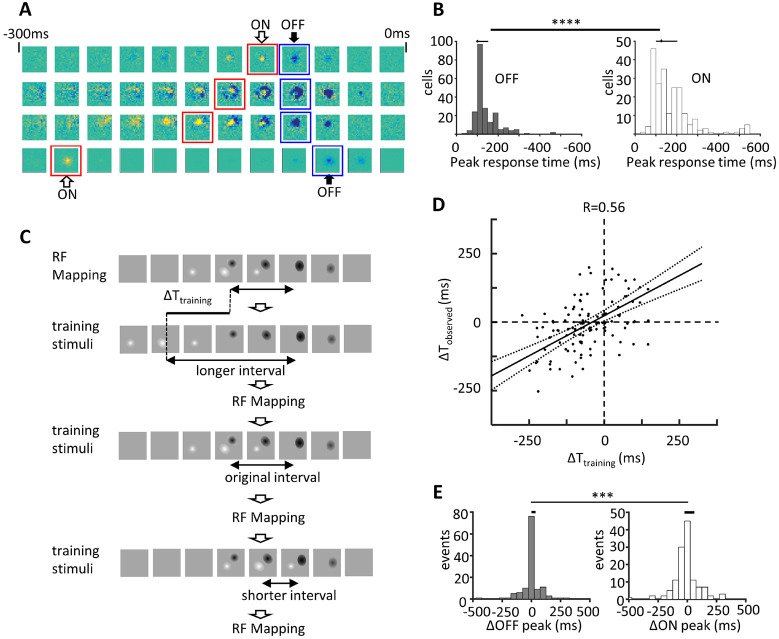
Natural visual scenes are dominated by sequences of transforming images. Spatial visual information is thought to be processed by detection of elemental stimulus features which are recomposed into scenes. How image information is integrated over time is unclear. We explored visual information encoding in the optic tectum. Unbiased stimulus presentation shows that the majority of tectal neurons recognize image sequences. This is achieved by temporally dynamic response properties, which encode complex image transitions over several hundred milliseconds. Calcium imaging reveals that neurons that encode spatiotemporal image sequences fire in spike sequences that predict a logical diagram of spatiotemporal information processing. Furthermore, the temporal scale of visual information is tuned by experience. This study indicates how neurons recognize dynamic visual scenes that transform over time.
Figures




References
-
- Hubel D. H., Wiesel T. N., Ferrier lecture. Functional architecture of macaque monkey visual cortex. Proc R Soc Lond B Biol Sci 198, 1–59 (1977). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources