

Systematic transcriptome profiling of hPSC-derived osteoblasts unveils CORIN's mastery in governing osteogenesis through CEBPD modulation
- PMID: 38925326
- PMCID: PMC11301355
- DOI: 10.1016/j.jbc.2024.107494
Systematic transcriptome profiling of hPSC-derived osteoblasts unveils CORIN's mastery in governing osteogenesis through CEBPD modulation
Abstract
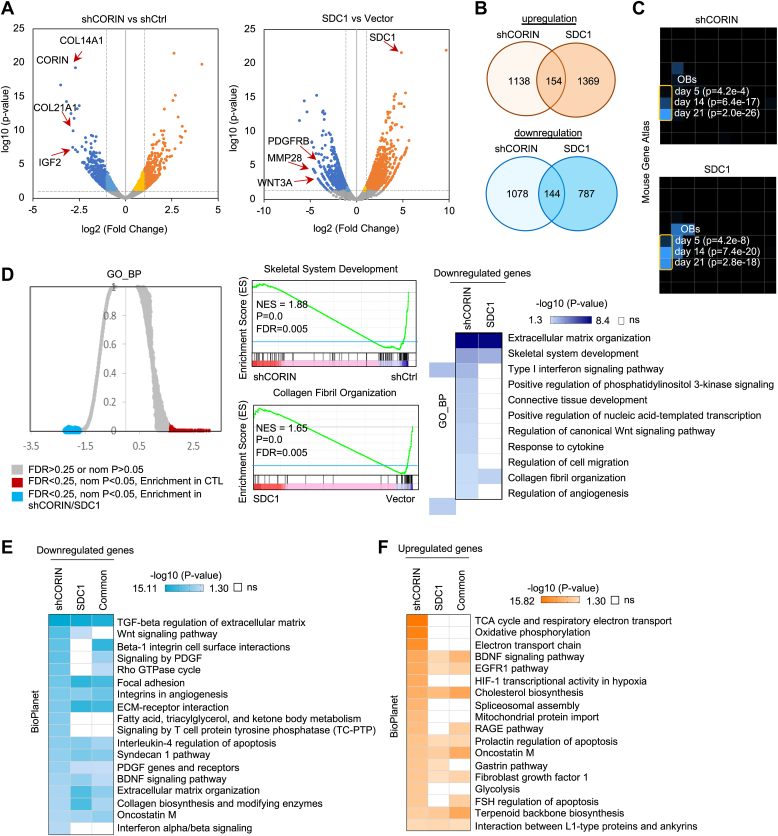
The commitment of stem cells to differentiate into osteoblasts is a highly regulated and complex process that involves the coordination of extrinsic signals and intrinsic transcriptional machinery. While rodent osteoblastic differentiation has been extensively studied, research on human osteogenesis has been limited by cell sources and existing models. Here, we systematically dissect human pluripotent stem cell-derived osteoblasts to identify functional membrane proteins and their downstream transcriptional networks involved in human osteogenesis. Our results reveal an enrichment of type II transmembrane serine protease CORIN in humans but not rodent osteoblasts. Functional analyses demonstrated that CORIN depletion significantly impairs osteogenesis. Genome-wide chromatin immunoprecipitation enrichment and mechanistic studies show that p38 MAPK-mediated CCAAT enhancer binding protein delta (CEBPD) upregulation is required for CORIN-modulated osteogenesis. Contrastingly, the type I transmembrane heparan sulfate proteoglycan SDC1 enriched in mesenchymal stem cells exerts a negative regulatory effect on osteogenesis through a similar mechanism. Chromatin immunoprecipitation-seq, bulk and single-cell transcriptomes, and functional validations indicated that CEBPD plays a critical role in controlling osteogenesis. In summary, our findings uncover previously unrecognized CORIN-mediated CEBPD transcriptomic networks in driving human osteoblast lineage commitment.
Keywords: ChIP-sequencing (ChIP-seq); induced pluripotent stem cells (iPSCs); mesenchymal stem cells (MSCs); osteoblasts; p38 MAPK.
Published by Elsevier Inc.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Mathews S., Bhonde R., Gupta P.K., Totey S. Extracellular matrix protein mediated regulation of the osteoblast differentiation of bone marrow derived human mesenchymal stem cells. Differentiation. 2012;84:185–192. - PubMed
-
- Neve A., Corrado A., Cantatore F.P. Osteoblast physiology in normal and pathological conditions. Cell Tissue Res. 2011;343:289–302. - PubMed
-
- Dong N., Niu Y., Chen Y., Sun S., Wu Q. Function and regulation of corin in physiology and disease. Biochem. Soc. Trans. 2020;48:1905–1916. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
