

Tumor cell-released autophagosomes (TRAPs) induce PD-L1-decorated NETs that suppress T-cell function to promote breast cancer pulmonary metastasis
- PMID: 38926151
- PMCID: PMC11216055
- DOI: 10.1136/jitc-2024-009082
Tumor cell-released autophagosomes (TRAPs) induce PD-L1-decorated NETs that suppress T-cell function to promote breast cancer pulmonary metastasis
Abstract
Background: Lung metastasis is the primary cause of breast cancer-related mortality. Neutrophil extracellular traps (NETs) are involved in the progression of breast cancer. However, the mechanism of NET formation is not fully understood. This study posits that tumor cell-released autophagosomes (TRAPs) play a crucial role in this process.
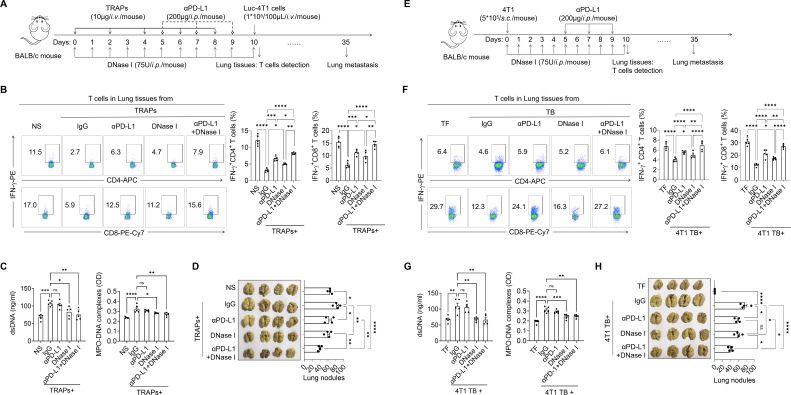
Methods: TRAPs were isolated from breast cancer cell lines to analyze their impact on NET formation in both human and mouse neutrophils. The study used both in vitro and in vivo models, including Toll-like receptor 4 (TLR4-/-) mice and engineered breast cancer cell lines. Immunofluorescence, ELISA, Western blotting, RNA sequencing, and flow cytometry were employed to dissect the signaling pathways leading to NET production and to explore their immunosuppressive effects, particularly focusing on the impact of NETs on T-cell function. The therapeutic potential of targeting TRAP-induced NETs and their immunosuppressive functions was evaluated using DNase I and αPD-L1 antibodies. Clinical relevance was assessed by correlating circulating levels of TRAPs and NETs with lung metastasis in patients with breast cancer.
Results: This study showed that TRAPs induced the formation of NETs in both human and mouse neutrophils by using the high mobility group box 1 and activating the TLR4-Myd88-ERK/p38 signaling axis. More importantly, PD-L1 carried by TRAP-induced NETs inhibited T-cell function in vitro and in vivo, thereby contributing to the formation of lung premetastatic niche (PMN) immunosuppression. In contrast, Becn1 KD-4T1 breast tumors with decreased circulating TRAPs in vivo reduced the formation of NETs, which in turn attenuated the immunosuppressive effects in PMN and resulted in a reduction of breast cancer pulmonary metastasis in murine models. Moreover, treatment with αPD-L1 in combination with DNase I that degraded NETs restored T-cell function and significantly reduced tumor metastasis. TRAP levels in the peripheral blood positively correlated with NET levels and lung metastasis in patients with breast cancer.
Conclusions: Our results demonstrate a novel role of TRAPs in the formation of PD-L1-decorated NETs, which may provide a new strategy for early detection and treatment of pulmonary metastasis in patients with breast cancer.
Keywords: Breast Cancer; Immune modulatory; Neutrophil.
© Author(s) (or their employer(s)) 2024. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures






References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous