

Synaptic architecture of leg and wing premotor control networks in Drosophila
- PMID: 38926579
- PMCID: PMC11356479
- DOI: 10.1038/s41586-024-07600-z
Synaptic architecture of leg and wing premotor control networks in Drosophila
Abstract
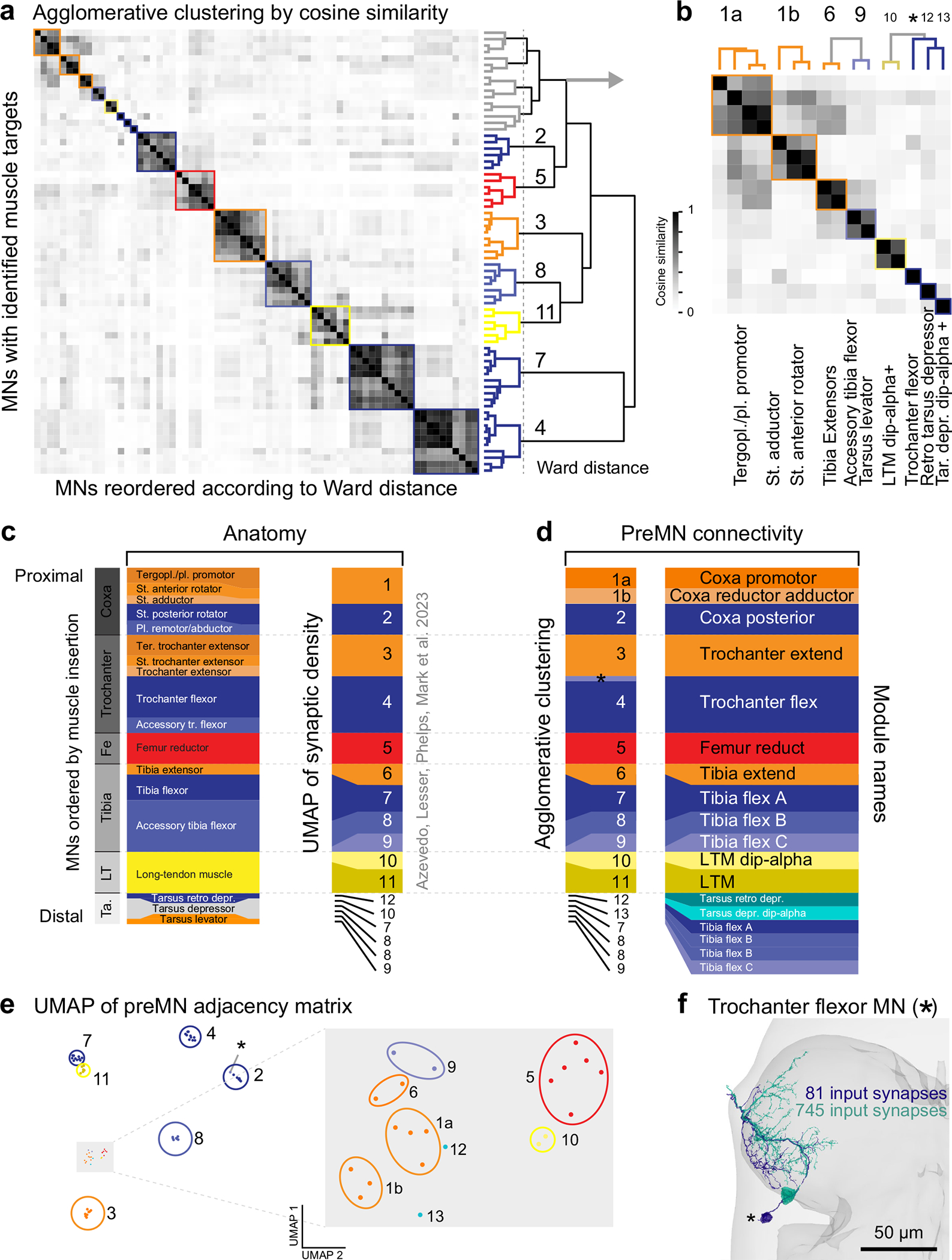
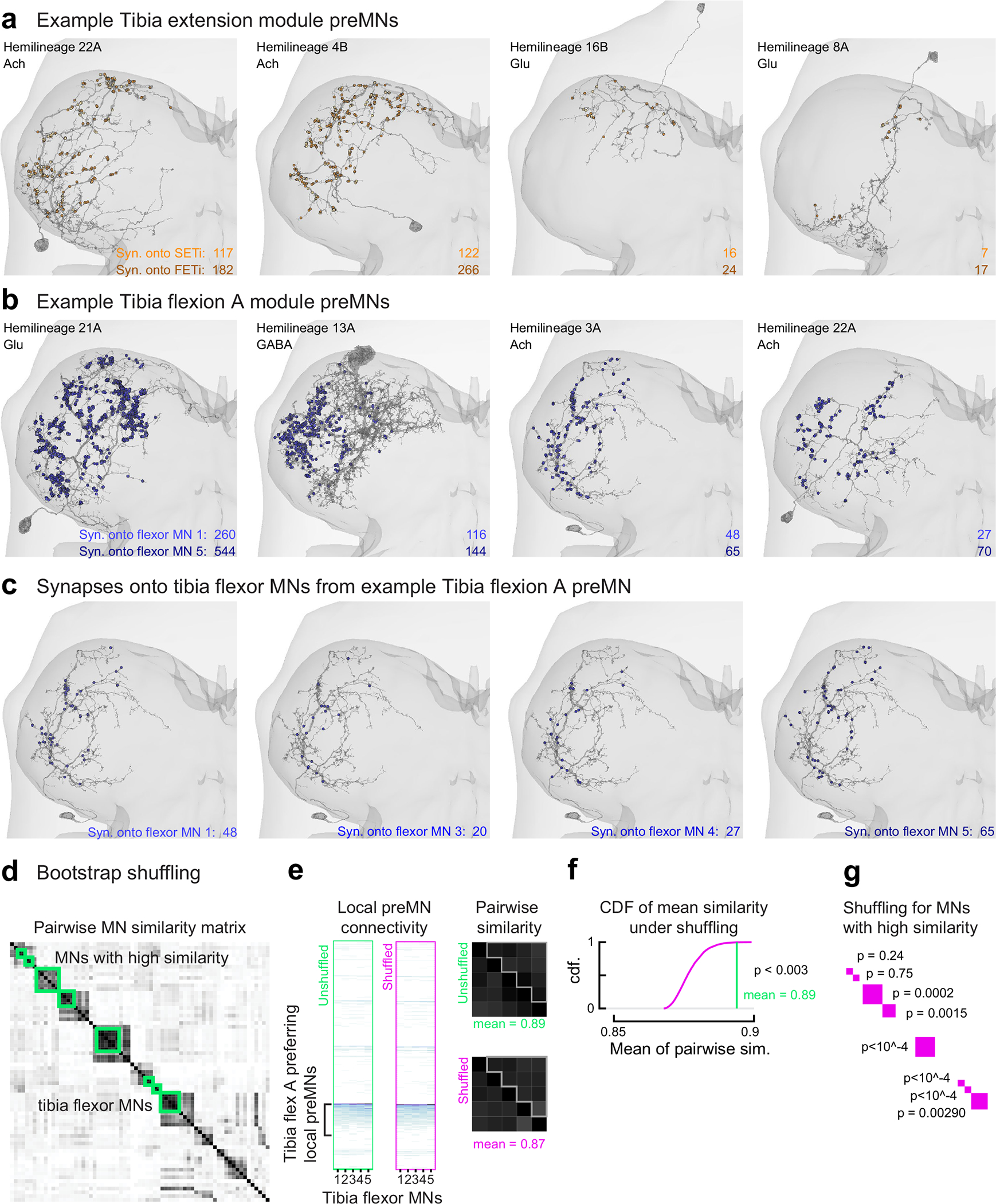
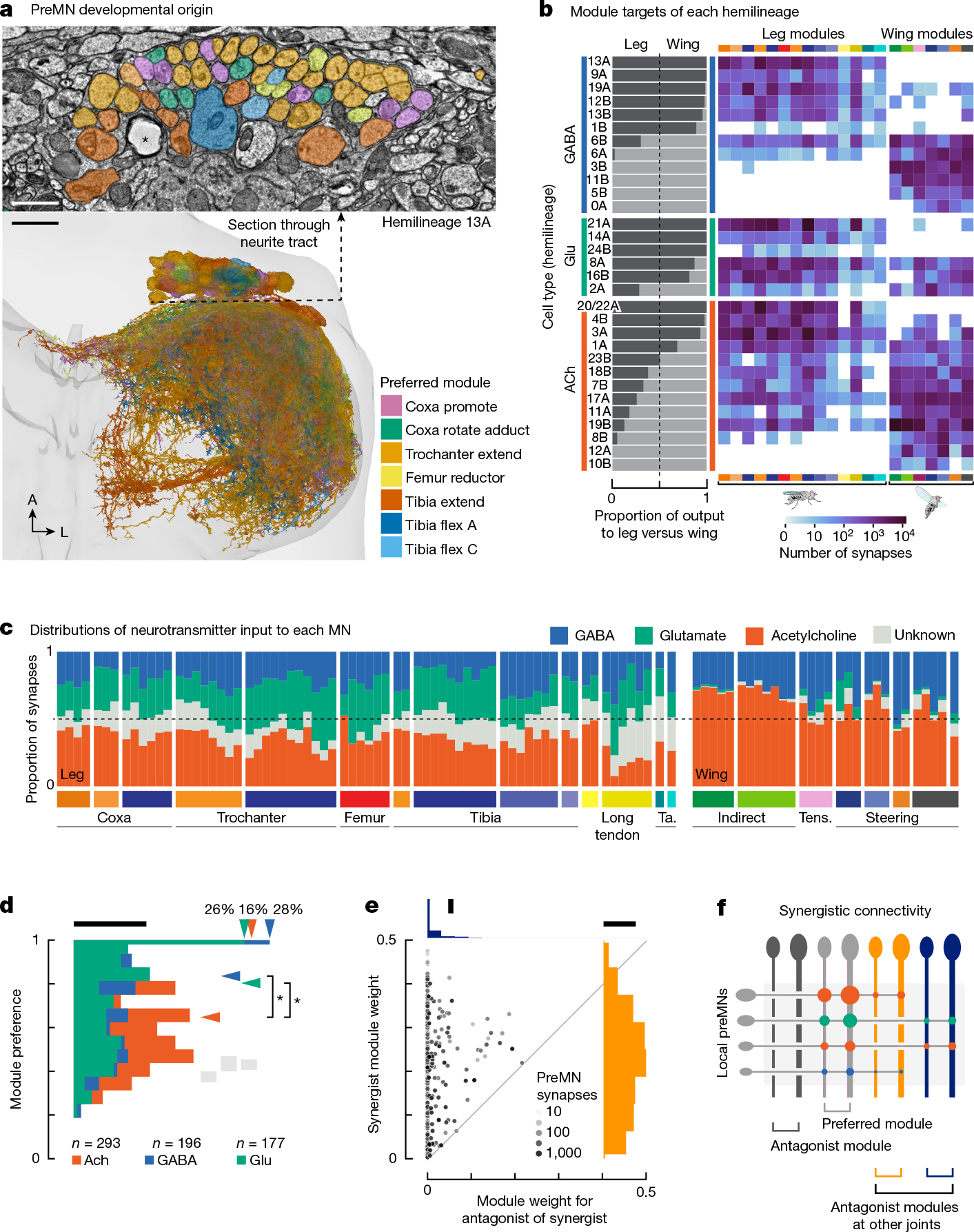
Animal movement is controlled by motor neurons (MNs), which project out of the central nervous system to activate muscles1. MN activity is coordinated by complex premotor networks that facilitate the contribution of individual muscles to many different behaviours2-6. Here we use connectomics7 to analyse the wiring logic of premotor circuits controlling the Drosophila leg and wing. We find that both premotor networks cluster into modules that link MNs innervating muscles with related functions. Within most leg motor modules, the synaptic weights of each premotor neuron are proportional to the size of their target MNs, establishing a circuit basis for hierarchical MN recruitment. By contrast, wing premotor networks lack proportional synaptic connectivity, which may enable more flexible recruitment of wing steering muscles. Through comparison of the architecture of distinct motor control systems within the same animal, we identify common principles of premotor network organization and specializations that reflect the unique biomechanical constraints and evolutionary origins of leg and wing motor control.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures









Update of
-
Synaptic architecture of leg and wing premotor control networks in Drosophila.bioRxiv [Preprint]. 2024 Apr 28:2023.05.30.542725. doi: 10.1101/2023.05.30.542725. bioRxiv. 2024. Update in: Nature. 2024 Jul;631(8020):369-377. doi: 10.1038/s41586-024-07600-z. PMID: 37398440 Free PMC article. Updated. Preprint.
Comment in
-
Neuroscience: A big step forward for motor control in Drosophila.Curr Biol. 2024 Sep 23;34(18):R859-R861. doi: 10.1016/j.cub.2024.07.097. Curr Biol. 2024. PMID: 39317156
References
-
- Kernell D The Motoneurone and Its Muscle Fibres (Oxford Univ. Press, 2006).
-
- Henneman E, Clamann HP, Gillies JD & Skinner RD Rank order of motoneurons within a pool: law of combination. J. Neurophysiol. 37, 1338–1349 (1974). - PubMed
-
- Tresch MC, Saltiel P, d’Avella A & Bizzi E Coordination and localization in spinal motor systems. Brain Res. Rev. 40, 66–79 (2002). - PubMed
-
- Ting LH & Macpherson JM A limited set of muscle synergies for force control during a postural task. J. Neurophysiol. 93, 609–613 (2005). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
