

ONECUT2 acts as a lineage plasticity driver in adenocarcinoma as well as neuroendocrine variants of prostate cancer
- PMID: 38932701
- PMCID: PMC11260453
- DOI: 10.1093/nar/gkae547
ONECUT2 acts as a lineage plasticity driver in adenocarcinoma as well as neuroendocrine variants of prostate cancer
Abstract
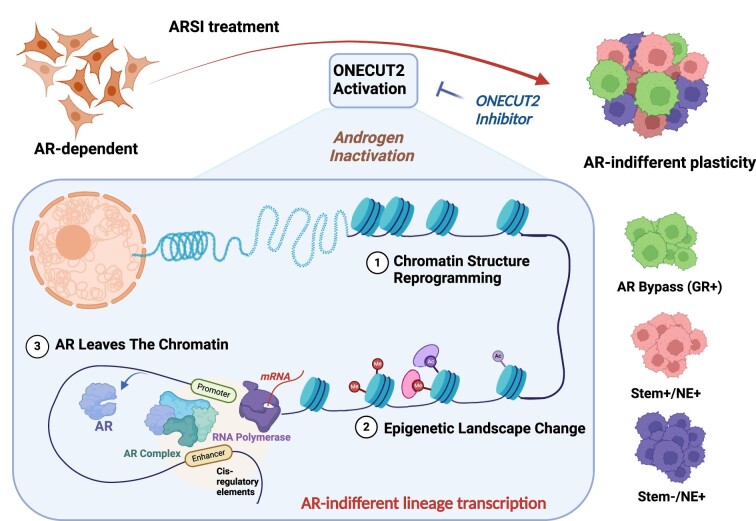
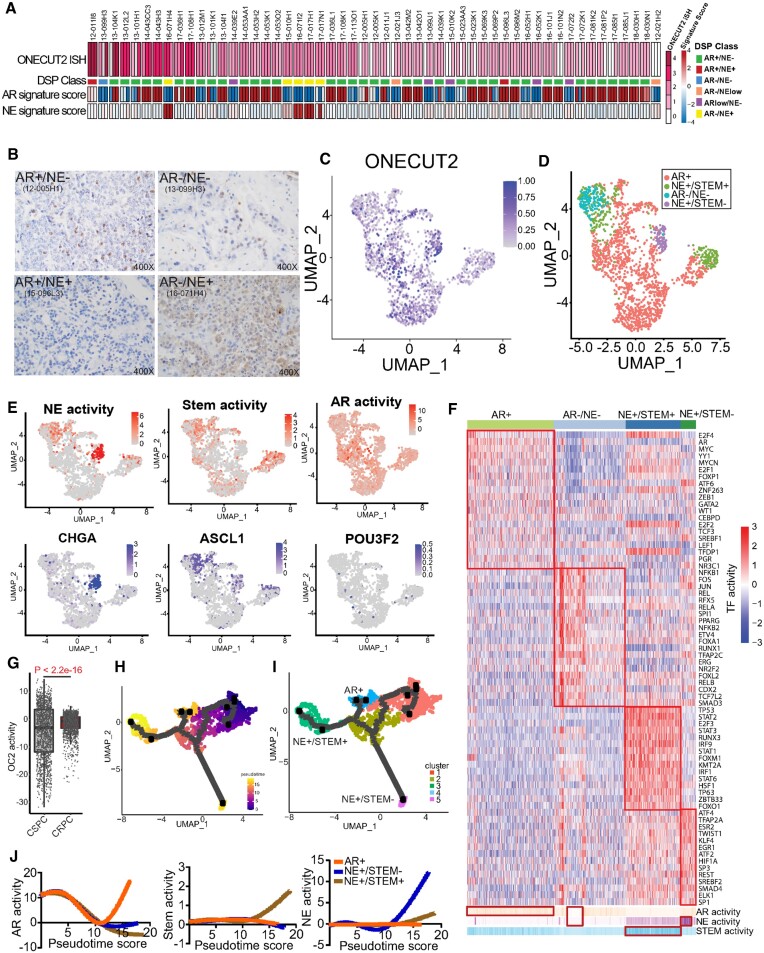
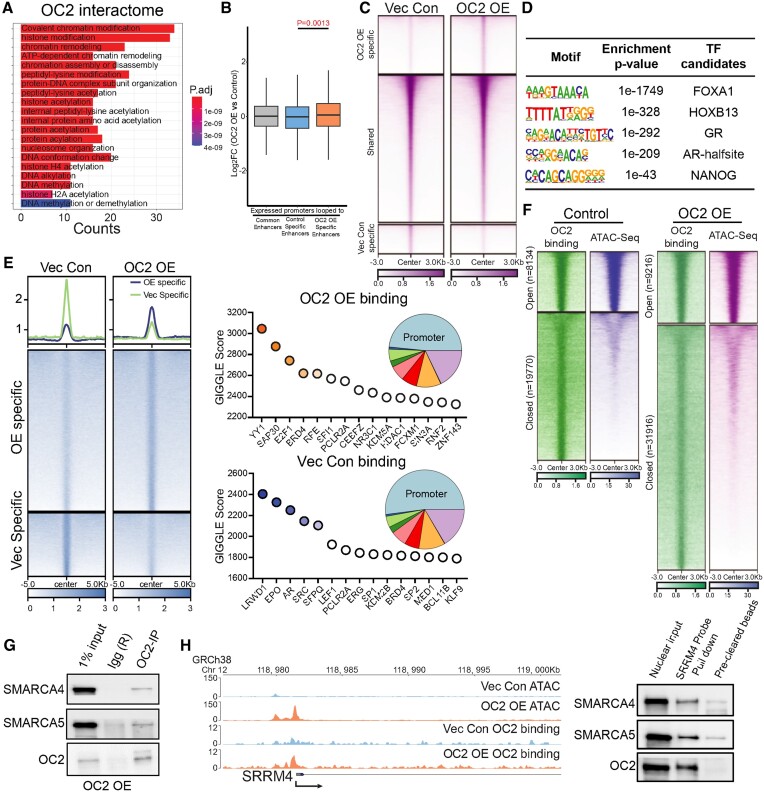
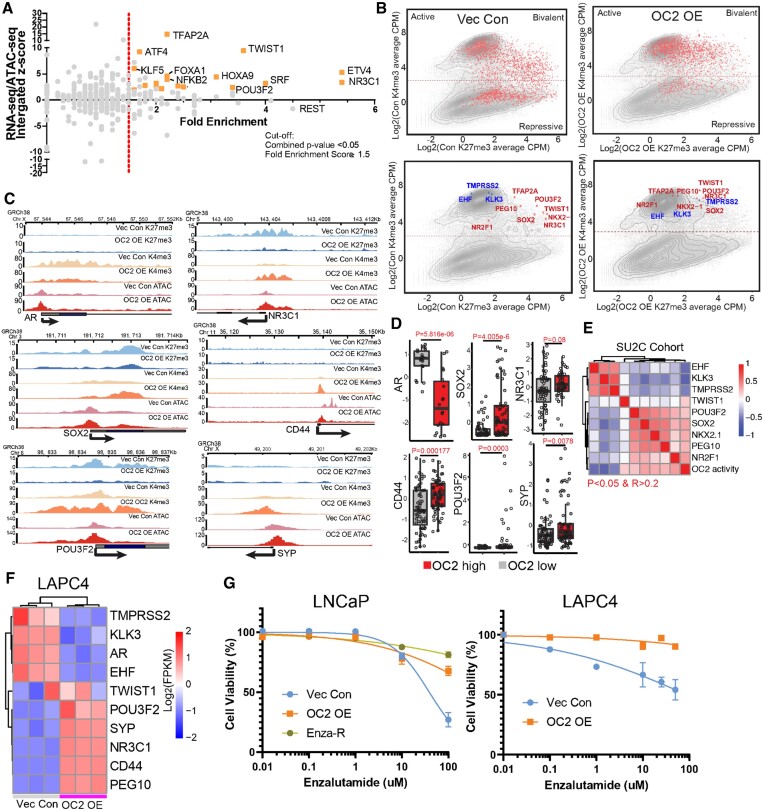
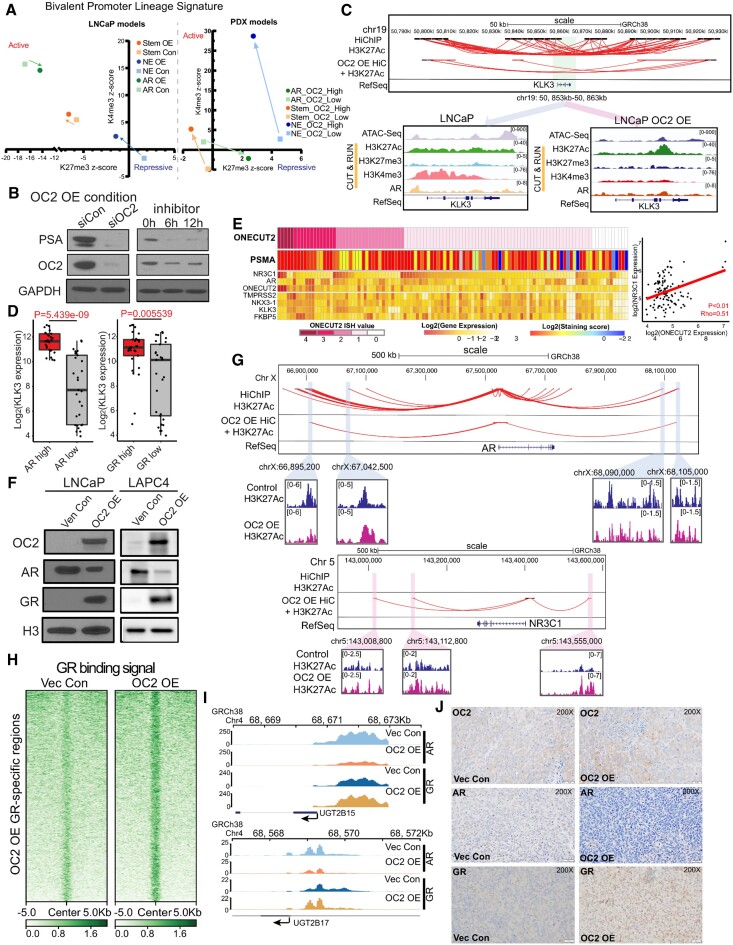
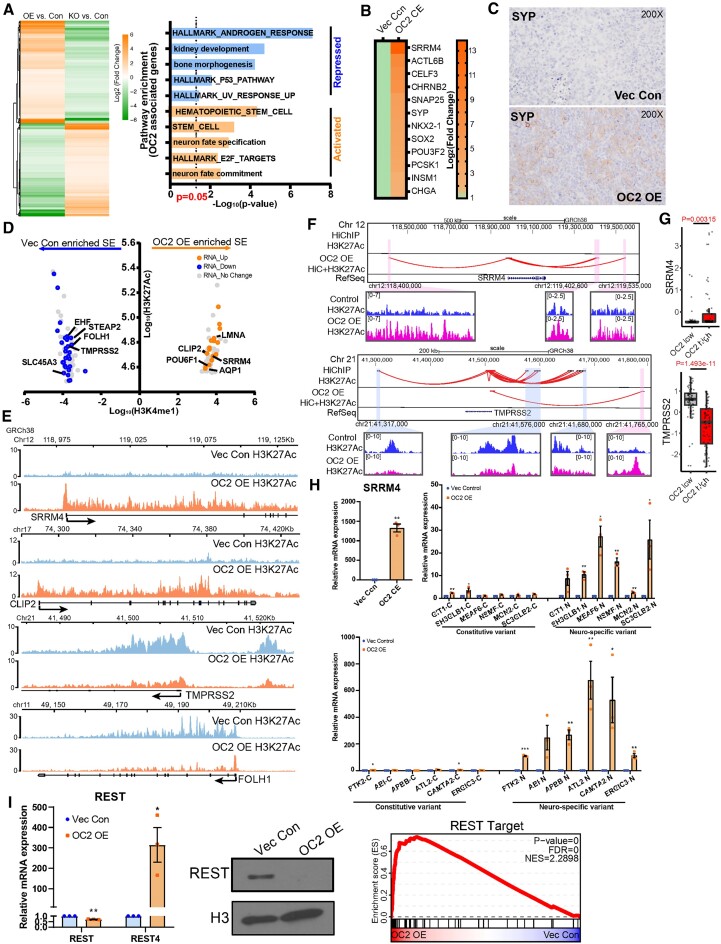
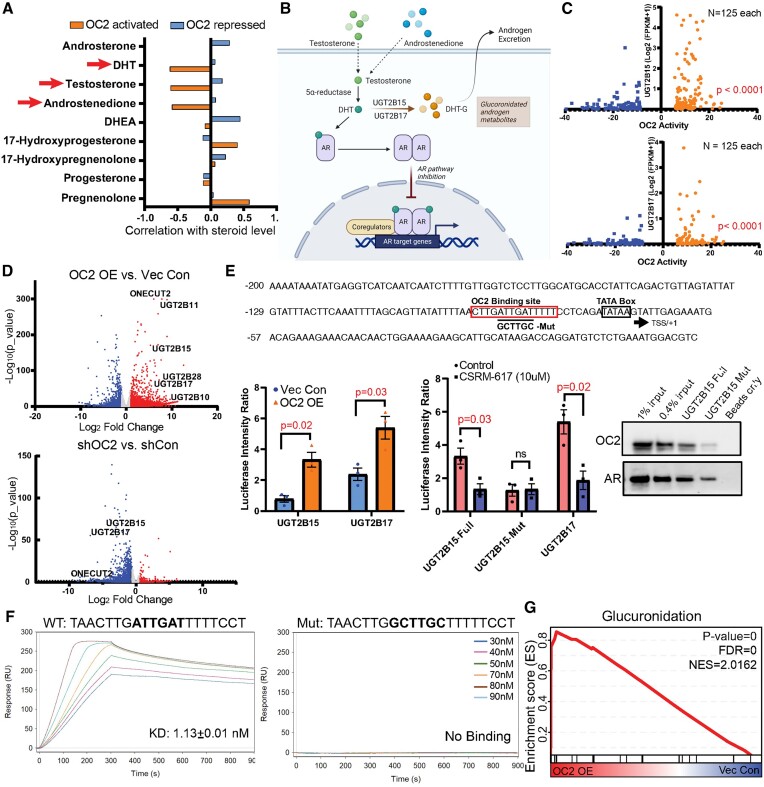
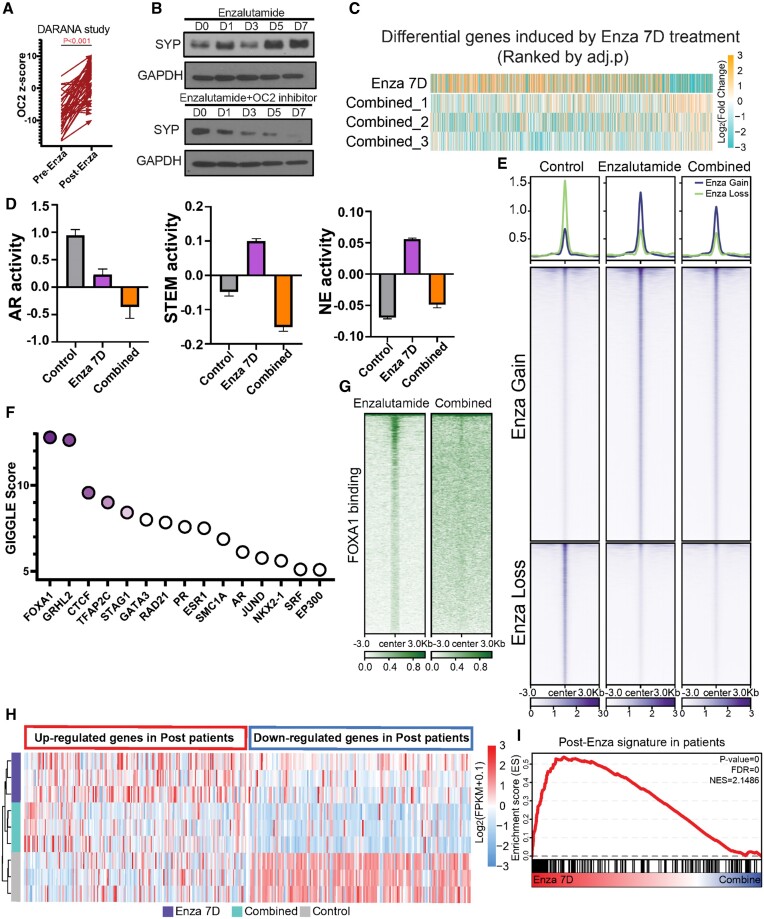
Androgen receptor- (AR-) indifference is a mechanism of resistance to hormonal therapy in prostate cancer (PC). Here we demonstrate that ONECUT2 (OC2) activates resistance through multiple drivers associated with adenocarcinoma, stem-like and neuroendocrine (NE) variants. Direct OC2 gene targets include the glucocorticoid receptor (GR; NR3C1) and the NE splicing factor SRRM4, which are key drivers of lineage plasticity. Thus, OC2, despite its previously described NEPC driver function, can indirectly activate a portion of the AR cistrome through epigenetic activation of GR. Mechanisms by which OC2 regulates gene expression include promoter binding, enhancement of genome-wide chromatin accessibility, and super-enhancer reprogramming. Pharmacologic inhibition of OC2 suppresses lineage plasticity reprogramming induced by the AR signaling inhibitor enzalutamide. These results demonstrate that OC2 activation promotes a range of drug resistance mechanisms associated with treatment-emergent lineage variation in PC and support enhanced efforts to therapeutically target OC2 as a means of suppressing treatment-resistant disease.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
