

Proximity Labeling Proteomics Reveals Kv1.3 Potassium Channel Immune Interactors in Microglia
- PMID: 38936775
- PMCID: PMC11780389
- DOI: 10.1016/j.mcpro.2024.100809
Proximity Labeling Proteomics Reveals Kv1.3 Potassium Channel Immune Interactors in Microglia
Abstract
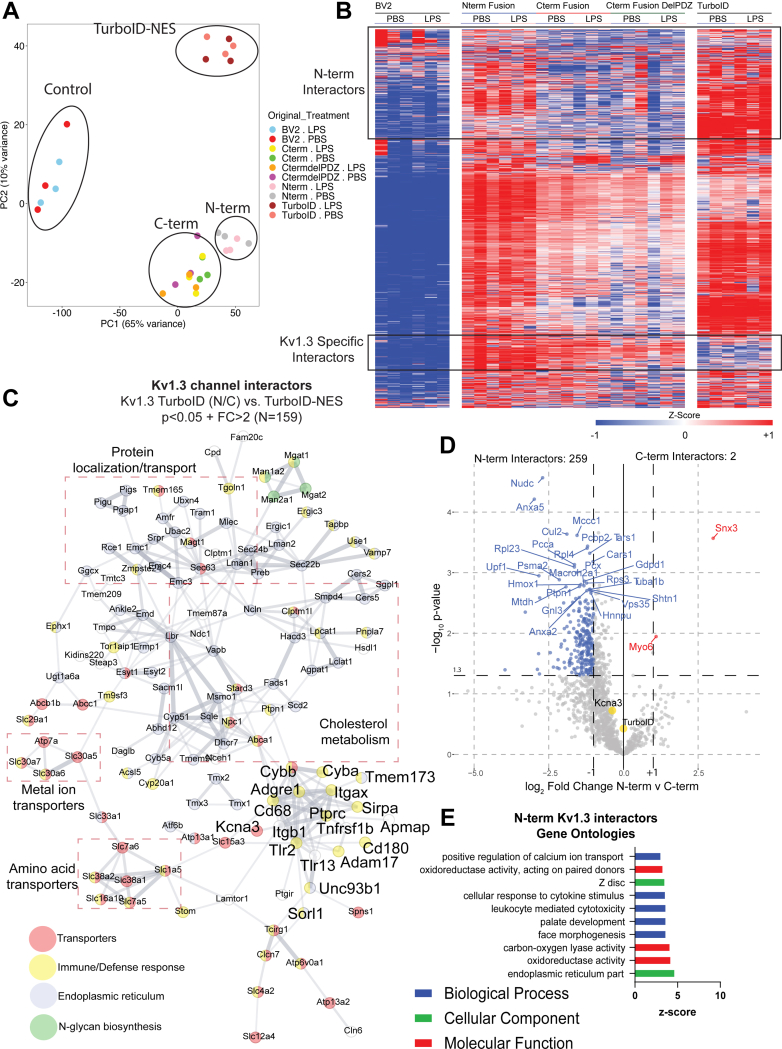
Microglia are resident immune cells of the brain and regulate its inflammatory state. In neurodegenerative diseases, microglia transition from a homeostatic state to a state referred to as disease-associated microglia (DAM). DAM express higher levels of proinflammatory signaling molecules, like STAT1 and TLR2, and show transitions in mitochondrial activity toward a more glycolytic response. Inhibition of Kv1.3 decreases the proinflammatory signature of DAM, though how Kv1.3 influences the response is unknown. Our goal was to identify the potential proteins interacting with Kv1.3 during transition to DAM. We utilized TurboID, a biotin ligase, fused to Kv1.3 to evaluate potential interacting proteins with Kv1.3 via mass spectrometry in BV-2 microglia following TLR4-mediated activation. Electrophysiology, Western blotting, and flow cytometry were used to evaluate Kv1.3 channel presence and TurboID biotinylation activity. We hypothesized that Kv1.3 contains domain-specific interactors that vary during a TLR4-induced inflammatory response, some of which are dependent on the PDZ-binding domain on the C terminus. We determined that the N terminus of Kv1.3 is responsible for trafficking Kv1.3 to the cell surface and mitochondria (e.g., NUDC, TIMM50). Whereas, the C terminus interacts with immune signaling proteins in a lipopolysaccharide-induced inflammatory response (e.g., STAT1, TLR2, and C3). There are 70 proteins that rely on the C-terminal PDZ-binding domain to interact with Kv1.3 (e.g., ND3, Snx3, and Sun1). Furthermore, we used Kv1.3 blockade to verify functional coupling between Kv1.3 and interferon-mediated STAT1 activation. Overall, we highlight that the Kv1.3 potassium channel functions beyond conducting the outward flux of potassium ions in an inflammatory context and that Kv1.3 modulates the activity of key immune signaling proteins, such as STAT1 and C3.
Keywords: Kv1.3 potassium channel; microglia; neurodegeneration; neuroinflammation; proximity labeling.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest H. W. is an inventor on a University of California patent claiming PAP-1 for immunosuppression. This patent has been abandoned because of its short remaining patent life. The authors declare no competing interests.
Figures







References
-
- Keren-Shaul H., Spinrad A., Weiner A., Matcovitch-Natan O., Dvir-Szternfeld R., Ulland T.K., et al. A unique microglia type associated with restricting development of Alzheimer’s disease. Cell. 2017;169:1276–1290.e17. - PubMed
-
- Deczkowska A., Keren-Shaul H., Weiner A., Colonna M., Schwartz M., Amit I. Disease-associated microglia: a universal immune sensor of neurodegeneration. Cell. 2018;173:1073–1081. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
