

TGR5 deficiency in excitatory neurons ameliorates Alzheimer's pathology by regulating APP processing
- PMID: 38941459
- PMCID: PMC11212731
- DOI: 10.1126/sciadv.ado1855
TGR5 deficiency in excitatory neurons ameliorates Alzheimer's pathology by regulating APP processing
Abstract
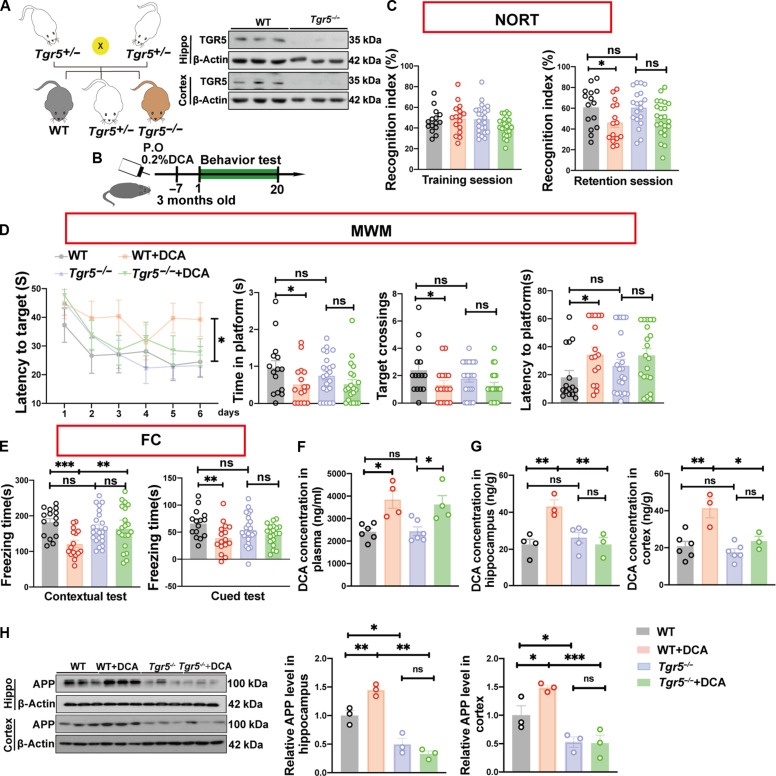
Bile acids (BAs) metabolism has a significant impact on the pathogenesis of Alzheimer's disease (AD). We found that deoxycholic acid (DCA) increased in brains of AD mice at an early stage. The enhanced production of DCA induces the up-regulation of the bile acid receptor Takeda G protein-coupled receptor (TGR5), which is also specifically increased in neurons of AD mouse brains at an early stage. The accumulation of exogenous DCA impairs cognitive function in wild-type mice, but not in TGR5 knockout mice. This suggests that TGR5 is the primary receptor mediating these effects of DCA. Furthermore, excitatory neuron-specific knockout of TGR5 ameliorates Aβ pathology and cognition impairments in AD mice. The underlying mechanism linking TGR5 and AD pathology relies on the downstream effectors of TGR5 and the APP production, which is succinctly concluded as a "p-STAT3-APH1-γ-secretase" signaling pathway. Our studies identified the critical role of TGR5 in the pathological development of AD.
Figures





References
-
- Jia W., Rajani C., Kaddurah-Daouk R., Li H., Expert insights: The potential role of the gut microbiome-bile acid-brain axis in the development and progression of Alzheimer's disease and hepatic encephalopathy. Med. Res. Rev. 40, 1496–1507 (2020). - PubMed
-
- Mahajan U. V., Varma V. R., Griswold M. E., Blackshear C. T., An Y., Oommen A. M., Varma S., Troncoso J. C., Pletnikova O., O'Brien R., Hohman T. J., Legido-Quigley C., Thambisetty M., Dysregulation of multiple metabolic networks related to brain transmethylation and polyamine pathways in Alzheimer disease: A targeted metabolomic and transcriptomic study. PLOS Med. 17, e1003012 (2020). - PMC - PubMed
-
- van Lengerich B., Zhan L., Xia D., Chan D., Joy D., Park J. I., Tatarakis D., Calvert M., Hummel S., Lianoglou S., Pizzo M. E., Prorok R., Thomsen E., Bartos L. M., Beumers P., Capell A., Davis S. S., de Weerd L., Dugas J. C., Duque J., Earr T., Gadkar K., Giese T., Gill A., Gnörich J., Ha C., Kannuswamy M., Kim D. J., Kunte S. T., Kunze L. H., Lac D., Lechtenberg K., Leung A. W., Liang C. C., Lopez I., McQuade P., Modi A., Torres V. O., Nguyen H. N., Pesämaa I., Propson N., Reich M., Robles-Colmenares Y., Schlepckow K., Slemann L., Solanoy H., Suh J. H., Thorne R. G., Vieira C., Wind-Mark K., Xiong K., Zuchero Y. J. Y., Diaz D., Dennis M. S., Huang F., Scearce-Levie K., Watts R. J., Haass C., Lewcock J. W., Di Paolo G., Brendel M., Sanchez P. E., Monroe K. M., A TREM2-activating antibody with a blood-brain barrier transport vehicle enhances microglial metabolism in Alzheimer's disease models. Nat. Neurosci. 26, 416–429 (2023). - PMC - PubMed
-
- Andersen J. V., Schousboe A., Verkhratsky A., Astrocyte energy and neurotransmitter metabolism in Alzheimer's disease: Integration of the glutamate/GABA-glutamine cycle. Prog. Neurobiol. 217, 102331 (2022). - PubMed
-
- Wang H., Tan Y. Z., Mu R. H., Tang S. S., Liu X., Xing S. Y., Long Y., Yuan D. H., Hong H., Takeda G protein-coupled receptor 5 modulates depression-like behaviors via hippocampal CA3 pyramidal neurons afferent to dorsolateral septum. Biol. Psychiatry 89, 1084–1095 (2021). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
