

Th2-biased immune responses to body migrating Ascaris larvae in primary infection are associated with pathology but not protection
- PMID: 38942904
- PMCID: PMC11213949
- DOI: 10.1038/s41598-024-65281-0
Th2-biased immune responses to body migrating Ascaris larvae in primary infection are associated with pathology but not protection
Abstract
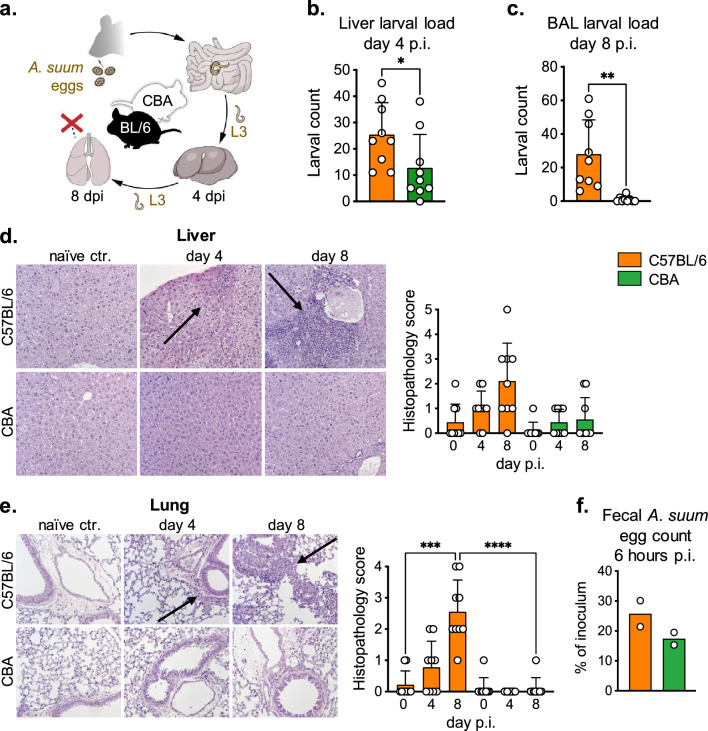
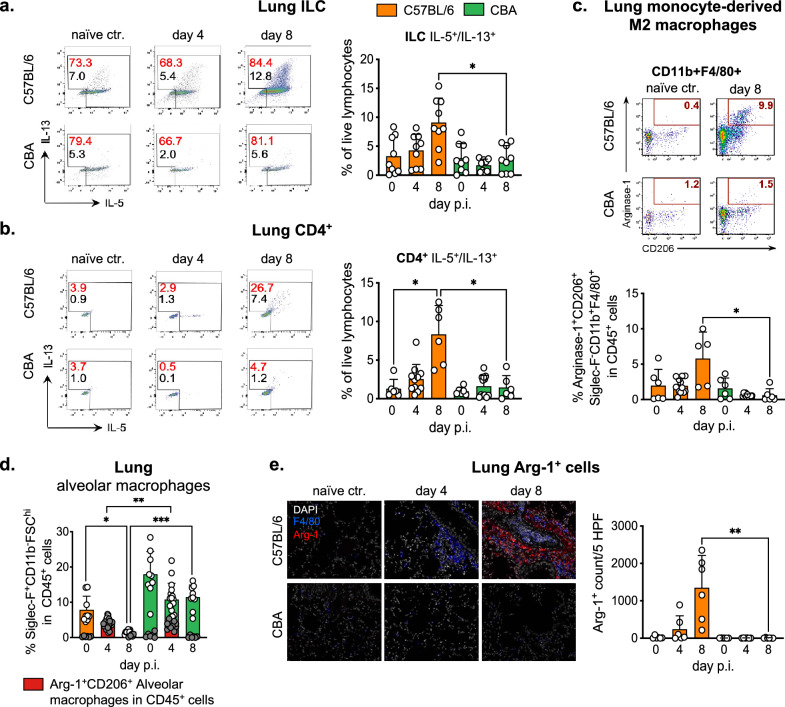
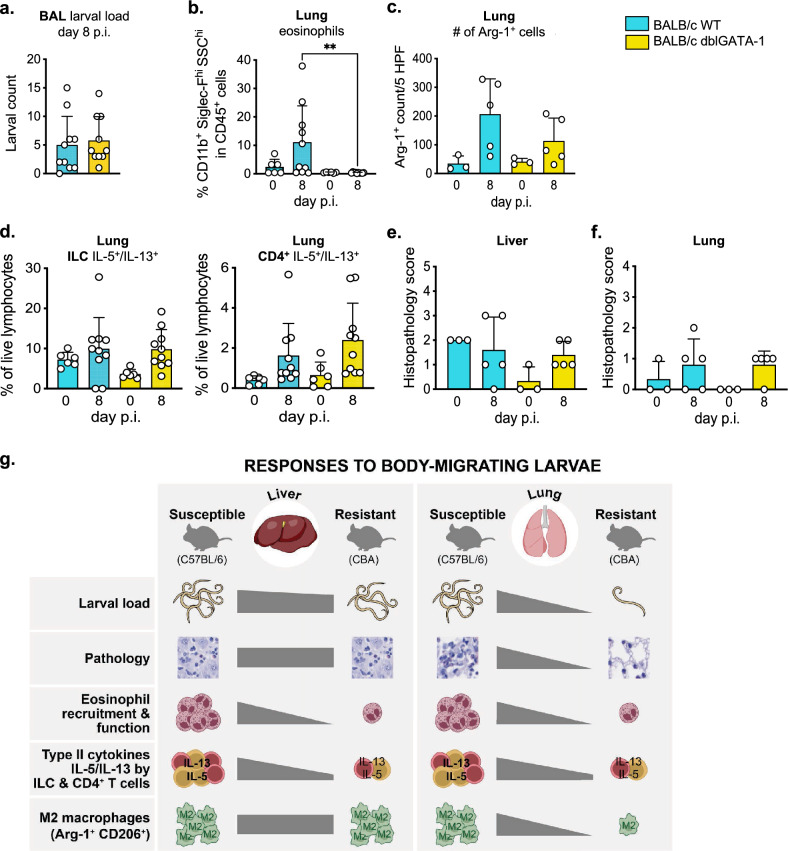
Helminth infections lead to an overdispersion of the parasites in humans as well as in animals. We asked whether early immune responses against migrating Ascaris larvae are responsible for the unequal distribution of worms in natural host populations and thus investigated a susceptible versus a resistant mouse strain. In mice, the roundworm larvae develop until the lung stage and thus early anti-Ascaris immune responses against the migrating larvae in the liver and lung can be deciphered. Our data show that susceptible C57BL/6 mice respond to Ascaris larval migration significantly stronger compared to resistant CBA mice and the anti-parasite reactivity is associated with pathology. Increased eosinophil recruitment was detected in the liver and lungs, but also in the spleen and peritoneal cavity of susceptible mice on day 8 post infection compared to resistant mice. In serum, eosinophil peroxidase levels were significantly higher only in the susceptible mice, indicating functional activity of the recruited eosinophils. This effect was associated with an increased IL-5/IL-13 production by innate lymphoid cells and CD4+ T cells and a pronounced type 2 macrophage polarization in the lungs of susceptible mice. Furthermore, a comparison of wildtype BALB/c and eosinophil-deficient dblGATA-1 BALB/c mice showed that eosinophils were not essential for the early control of migrating Ascaris larvae. In conclusion, in primary infection, a strong local and systemic type 2 immune response during hepato-tracheal helminth larval migration is associated with pathology rather than protection.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
