

A proposed pathway from D-glucose to D-arabinose in eukaryotes
- PMID: 38944124
- PMCID: PMC11301363
- DOI: 10.1016/j.jbc.2024.107500
A proposed pathway from D-glucose to D-arabinose in eukaryotes
Abstract
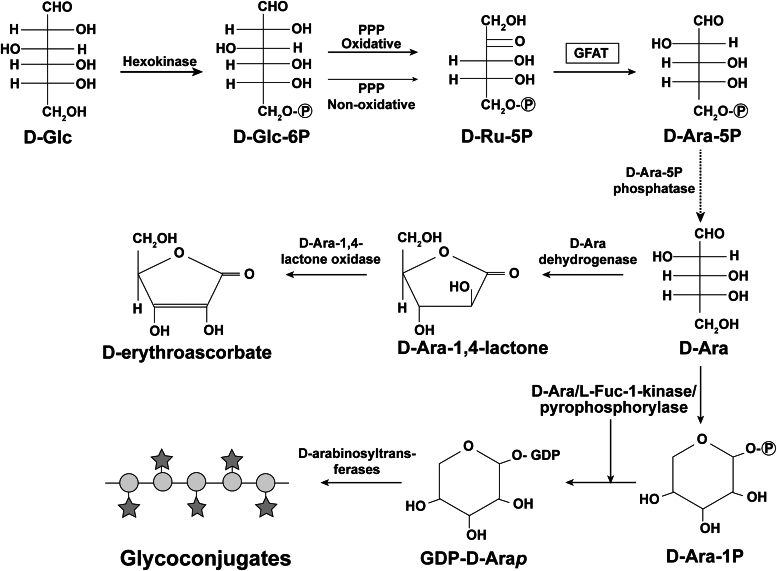
In eukaryotes, the D-enantiomer of arabinose (D-Ara) is an intermediate in the biosynthesis of D-erythroascorbate in yeast and fungi and in the biosynthesis of the nucleotide sugar GDP-α-D-arabinopyranose (GDP-D-Arap) and complex α-D-Arap-containing surface glycoconjugates in certain trypanosomatid parasites. Whereas the biosynthesis of D-Ara in prokaryotes is well understood, the route from D-glucose (D-Glc) to D-Ara in eukaryotes is unknown. In this paper, we study the conversion of D-Glc to D-Ara in the trypanosomatid Crithidia fasciculata using positionally labeled [13C]-D-Glc and [13C]-D-ribose ([13C]-D-Rib) precursors and a novel derivatization and gas chromatography-mass spectrometry procedure applied to a terminal metabolite, lipoarabinogalactan. These data implicate the both arms of pentose phosphate pathway and a likely role for D-ribulose-5-phosphate (D-Ru-5P) isomerization to D-Ara-5P. We tested all C. fasciculata putative sugar and polyol phosphate isomerase genes for their ability to complement a D-Ara-5P isomerase-deficient mutant of Escherichia coli and found that one, the glutamine fructose-6-phosphate aminotransferase (GFAT) of glucosamine biosynthesis, was able to rescue the E. coli mutant. We also found that GFAT genes of other trypanosomatid parasites, and those of yeast and human origin, could complement the E. coli mutant. Finally, we demonstrated biochemically that recombinant human GFAT can isomerize D-Ru-5P to D-Ara5P. From these data, we postulate a general eukaryotic pathway from D-Glc to D-Ara and discuss its possible significance. With respect to C. fasciculata, we propose that D-Ara is used not only for the synthesis of GDP-D-Arap and complex surface glycoconjugates but also in the synthesis of D-erythroascorbate.
Keywords: Crithidia fasciculata; D-arabinose; D-erythroascorbate; glucose metabolism; pentose phosphate pathway.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest Michael Ferguson reports financial support was provided by Wellcome Trust (Investigator Award 101842/Z13/Z). The authors declare that they have no conflicts of interest with the contents of this article.
Figures




References
-
- Huh W.K., Lee B.H., Kim S.T., Kim Y.R., Rhie G.E., Baek Y.W., et al. D-Erythroascorbic acid is an important antioxidant molecule in Saccharomyces cerevisiae. Mol. Microbiol. 1998;30:895–903. - PubMed
-
- Baroja-Mazo A., del Valle P., Rua J., de Cima S., Busto F., de Arriaga D., et al. Characterisation and biosynthesis of D-erythroascorbic acid in Phycomyces blakesleeanus. Fungal Genet. Biol. 2005;42:390–402. - PubMed
-
- McConville M.J., Thomas-Oates J.E., Ferguson M.A., Homans S.W. Structure of the lipophosphoglycan from Leishmania major. J. Biol. Chem. 1990;265:19611–19623. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
