

This is a preprint.
Large-Scale Genome-Wide Optimization and Prediction of the Cre Recombinase System for Precise Genome Manipulation in Mice
- PMID: 38948742
- PMCID: PMC11212873
- DOI: 10.1101/2024.06.14.599022
Large-Scale Genome-Wide Optimization and Prediction of the Cre Recombinase System for Precise Genome Manipulation in Mice
Update in
-
Systematic optimization and prediction of cre recombinase for precise genome editing in mice.Genome Biol. 2025 Apr 4;26(1):85. doi: 10.1186/s13059-025-03560-3. Genome Biol. 2025. PMID: 40186303 Free PMC article.
Abstract
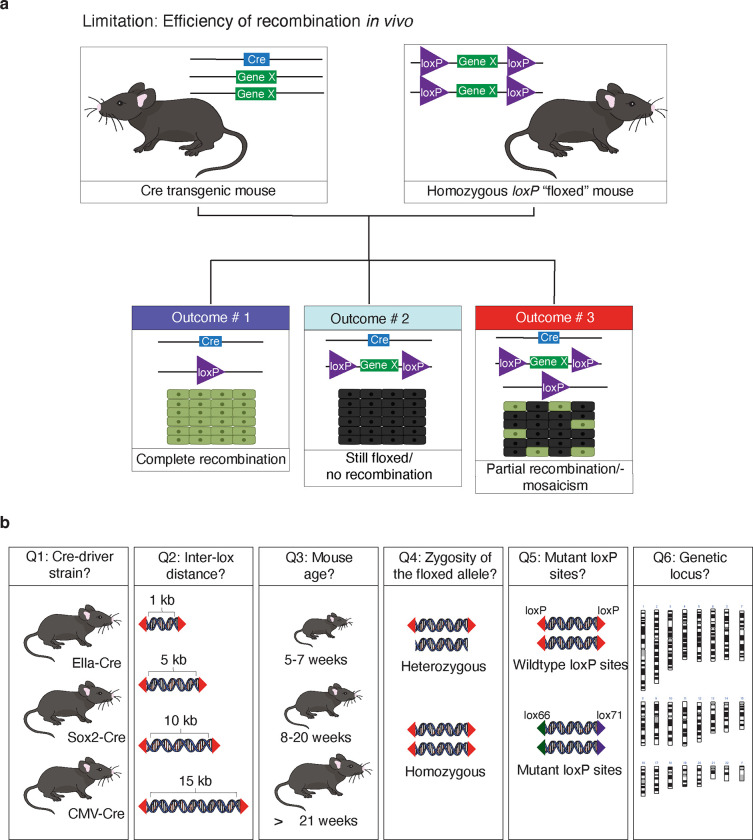
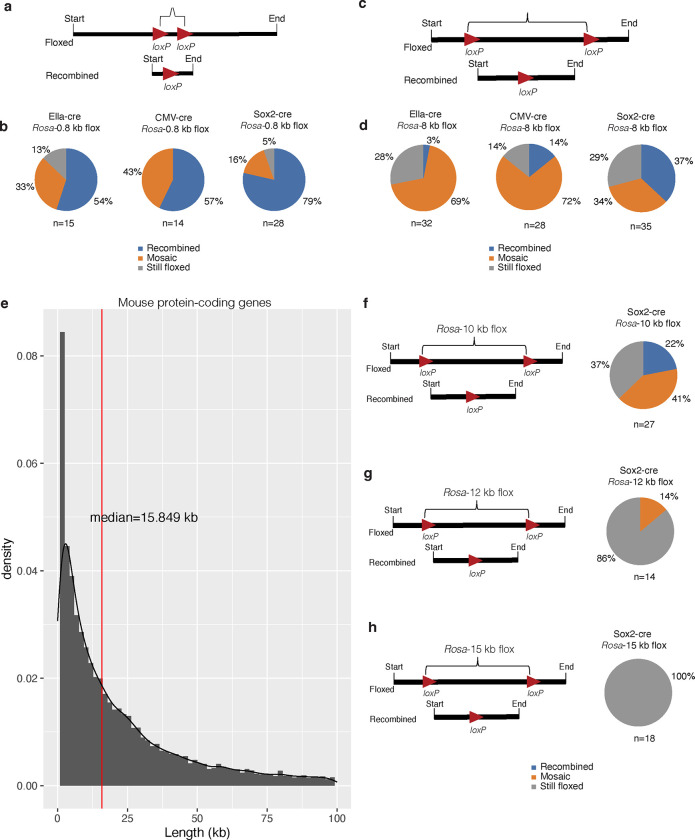
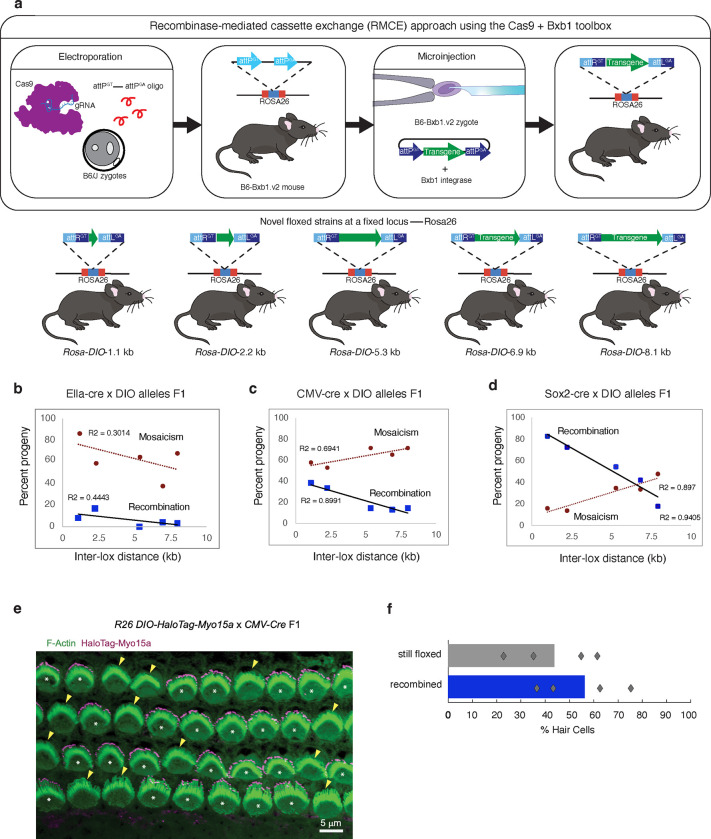
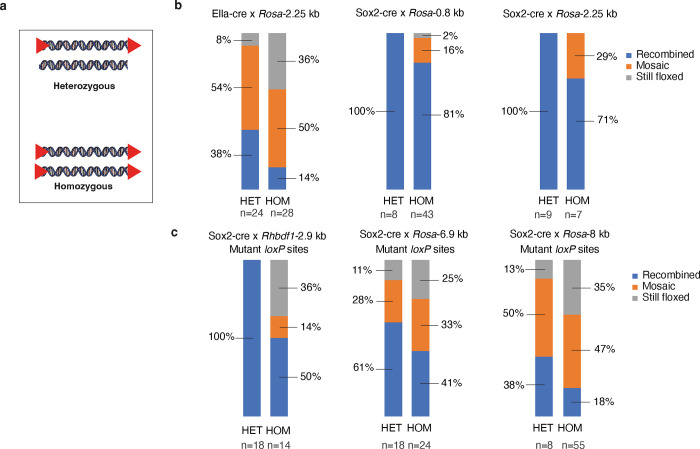
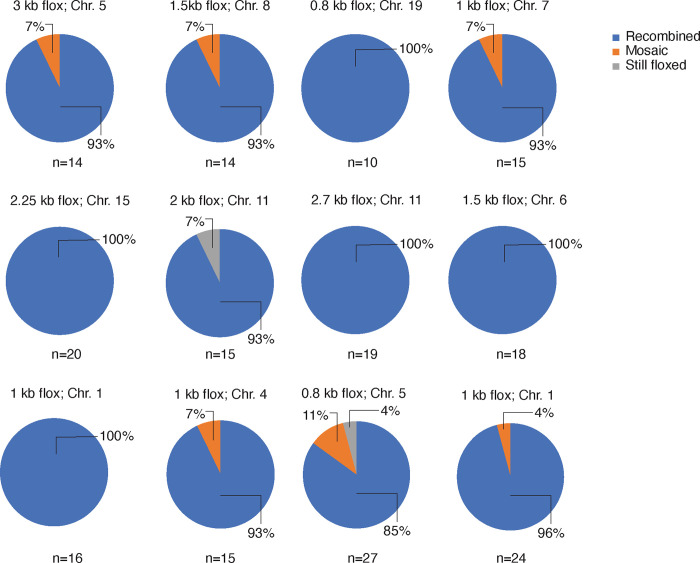
The Cre-Lox recombination system is a powerful tool in mouse genetics, offering spatial-temporal control over gene expression and facilitating the large-scale generation of conditional knockout mice. Its versatility also extends to other research models, such as rats, pigs, and zebrafish. However, the Cre-Lox technology presents a set of challenges that includes high costs, a time-intensive process, and the occurrence of unpredictable recombination events, which can lead to unexpected phenotypic outcomes. To better understand factors affecting recombination, we embarked on a systematic and genome-wide analysis of Cre-mediated recombination in mice. To ensure uniformity and reproducibility, we generated 11 novel strains with conditional alleles at the ROSA26 locus, utilizing a single inbred mouse strain background, C57BL/6J. We examined several factors influencing Cre-recombination, including the inter-loxP distance, mutant loxP sites, the zygosity of the conditional alleles, chromosomal location, and the age of the breeders. We discovered that the selection of the Cre-driver strain profoundly impacts recombination efficiency. We also found that successful and complete recombination is best achieved when loxP sites are spaced between 1 to 4 kb apart, with mutant loxP sites facilitating recombination at distances of 1 to 3 kb. Furthermore, we demonstrate that complete recombination does not occur at an inter-loxP distance of ≥ 15 kb with wildtype loxP sites, nor at a distance of ≥ 7 kb with mutant lox71/66 sites. Interestingly, the age of the Cre-driver mouse at the time of breeding emerged as a critical factor in recombination efficiency, with best results observed between 8 and 20 weeks old. Moreover, crossing heterozygous floxed alleles with the Cre-driver strain resulted in more efficient recombination than using homozygous floxed alleles. Lastly, maintaining an inter-loxP distance of 4 kb or less ensures efficient recombination of the conditional allele, regardless of the chromosomal location. While CRISPR/Cas has revolutionized genome editing in mice, Cre-Lox technology remains a cornerstone for the generation of sophisticated alleles and for precise control of gene expression in mice. The knowledge gained here will enable investigators to select a Cre-Lox approach that is most efficient for their desired outcome in the generation of both germline and non-germline mouse models of human disease, thereby reducing time and cost of Cre-Lox technology-mediated genome modification.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources