

This is a preprint.
Modeling Diffusive Search by Non-Adaptive Sperm: Empirical and Computational Insights
- PMID: 38948799
- PMCID: PMC11212867
- DOI: 10.1101/2024.06.17.599386
Modeling Diffusive Search by Non-Adaptive Sperm: Empirical and Computational Insights
Update in
-
Modeling diffusive search by non-adaptive sperm: Empirical and computational insights.PLoS Comput Biol. 2025 Apr 17;21(4):e1012865. doi: 10.1371/journal.pcbi.1012865. eCollection 2025 Apr. PLoS Comput Biol. 2025. PMID: 40244975 Free PMC article.
Abstract
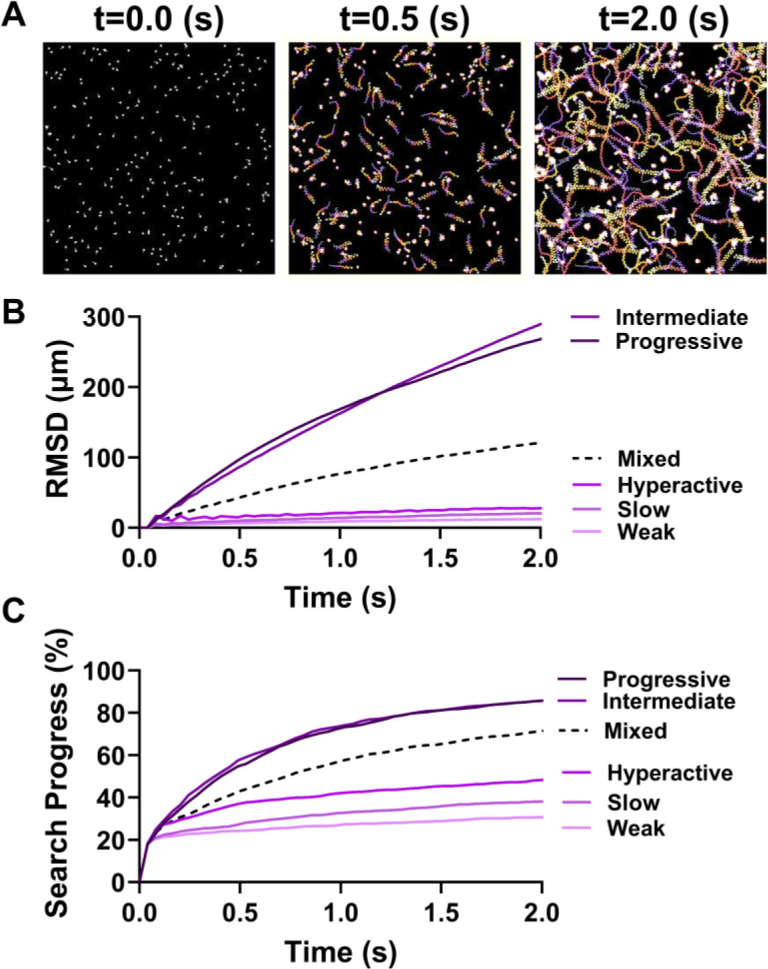
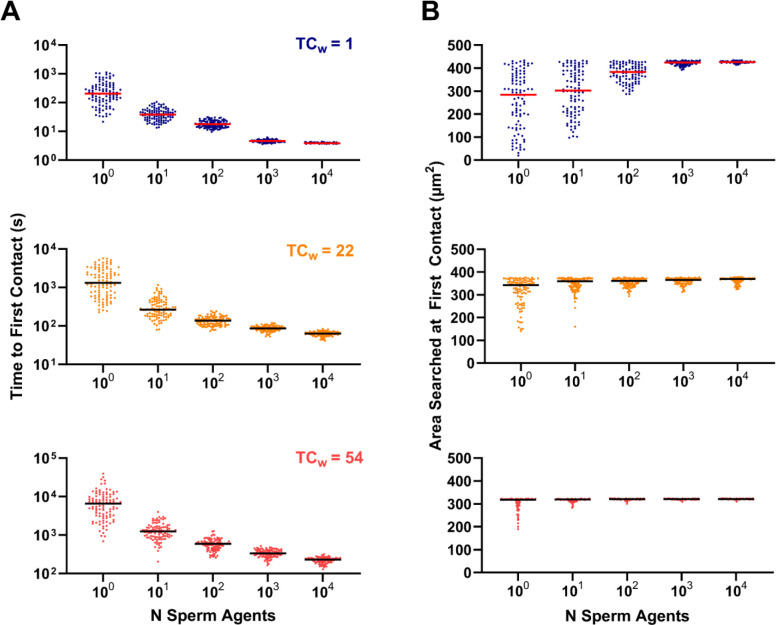
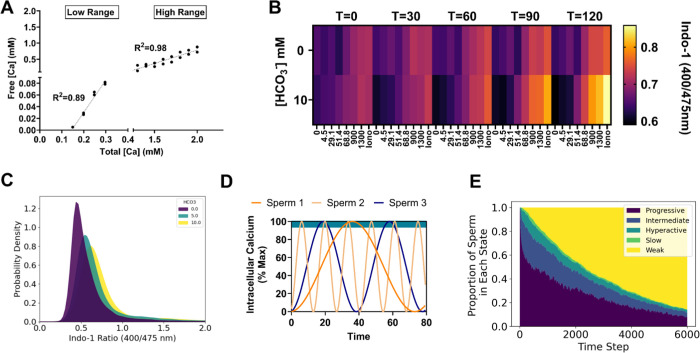
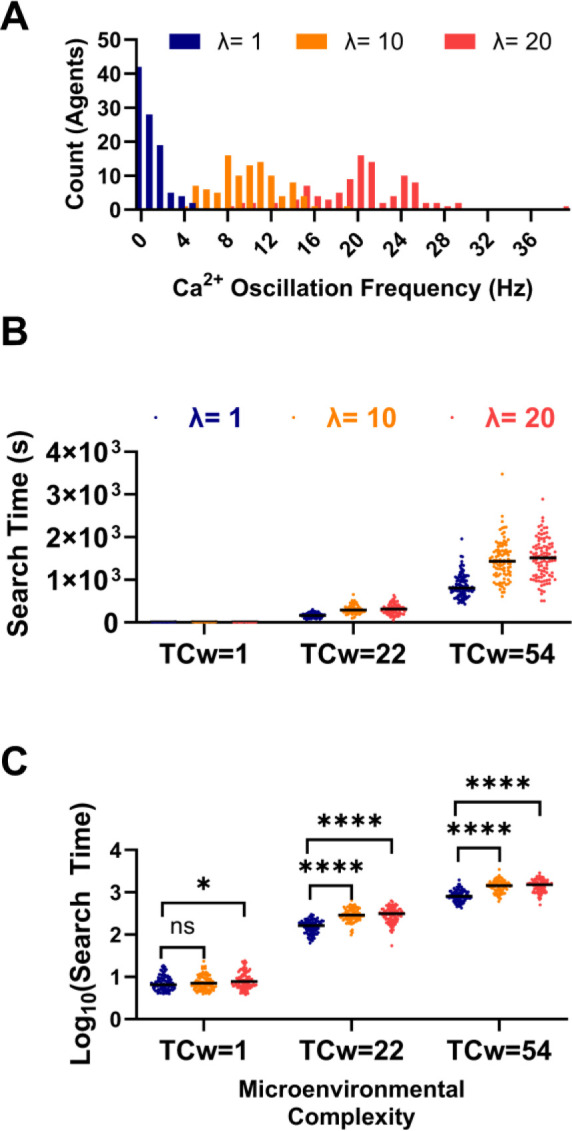
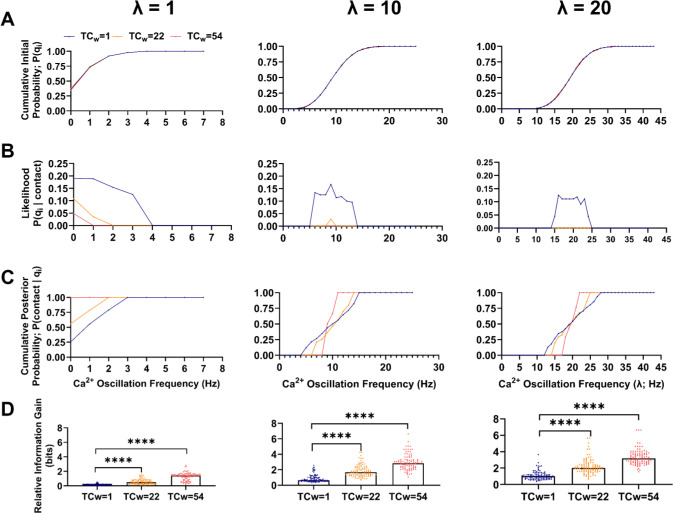
During fertilization, mammalian sperm undergo a winnowing selection process that reduces the candidate pool of potential fertilizers from ~106-1011 cells to 101-102 cells (depending on the species). Classical sperm competition theory addresses the positive or 'stabilizing' selection acting on sperm phenotypes within populations of organisms but does not strictly address the developmental consequences of sperm traits among individual organisms that are under purifying selection during fertilization. It is the latter that is of utmost concern for improving assisted reproductive technologies (ART) because 'low fitness' sperm may be inadvertently used for fertilization during interventions that rely heavily on artificial sperm selection, such as intracytoplasmic sperm injection (ICSI). Importantly, some form of sperm selection is used in nearly all forms of ART (e.g., differential centrifugation, swim-up, or hyaluronan binding assays, etc.). To date, there is no unifying quantitative framework (i.e., theory of sperm selection) that synthesizes causal mechanisms of selection with observed natural variation in individual sperm traits. In this report, we reframe the physiological function of sperm as a collective diffusive search process and develop multi-scale computational models to explore the causal dynamics that constrain sperm 'fitness' during fertilization. Several experimentally useful concepts are developed, including a probabilistic measure of sperm 'fitness' as well as an information theoretic measure of the magnitude of sperm selection, each of which are assessed under systematic increases in microenvironmental selective pressure acting on sperm motility patterns.
Conflict of interest statement
Disclosures The authors declare no conflicts of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources