

Primary cilia formation requires the Leigh syndrome-associated mitochondrial protein NDUFAF2
- PMID: 38949024
- PMCID: PMC11213510
- DOI: 10.1172/JCI175560
Primary cilia formation requires the Leigh syndrome-associated mitochondrial protein NDUFAF2
Abstract
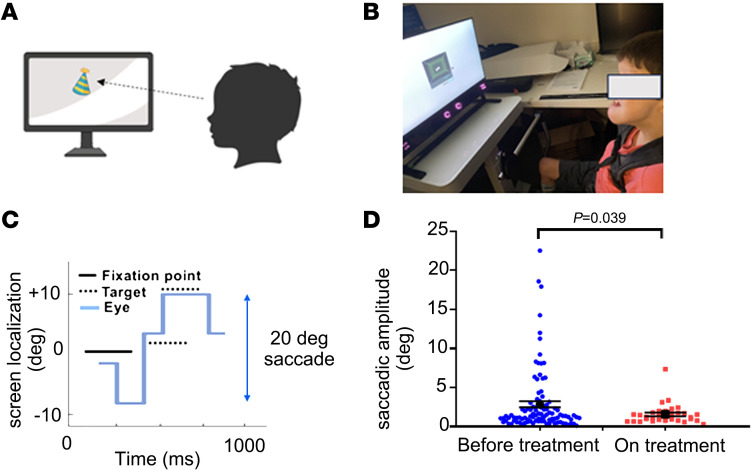
Mitochondria-related neurodegenerative diseases have been implicated in the disruption of primary cilia function. Mutation in an intrinsic mitochondrial complex I component NDUFAF2 has been identified in Leigh syndrome, a severe inherited mitochondriopathy. Mutations in ARMC9, which encodes a basal body protein, cause Joubert syndrome, a ciliopathy with defects in the brain, kidney, and eye. Here, we report a mechanistic link between mitochondria metabolism and primary cilia signaling. We discovered that loss of NDUFAF2 caused both mitochondrial and ciliary defects in vitro and in vivo and identified NDUFAF2 as a binding partner for ARMC9. We also found that NDUFAF2 was both necessary and sufficient for cilia formation and that exogenous expression of NDUFAF2 rescued the ciliary and mitochondrial defects observed in cells from patients with known ARMC9 deficiency. NAD+ supplementation restored mitochondrial and ciliary dysfunction in ARMC9-deficient cells and zebrafish and ameliorated the ocular motility and motor deficits of a patient with ARMC9 deficiency. The present results provide a compelling mechanistic link, supported by evidence from human studies, between primary cilia and mitochondrial signaling. Importantly, our findings have significant implications for the development of therapeutic approaches targeting ciliopathies.
Keywords: Cell biology; Neurodegeneration; Neurological disorders; Ophthalmology; Retinopathy.
Conflict of interest statement
Figures
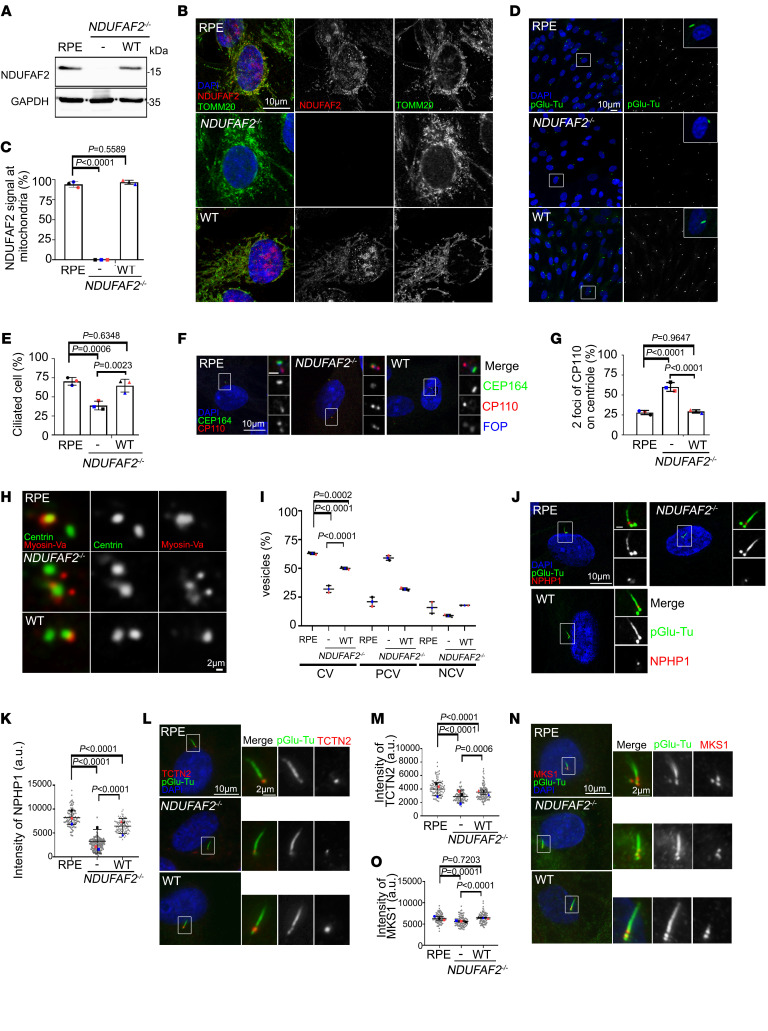

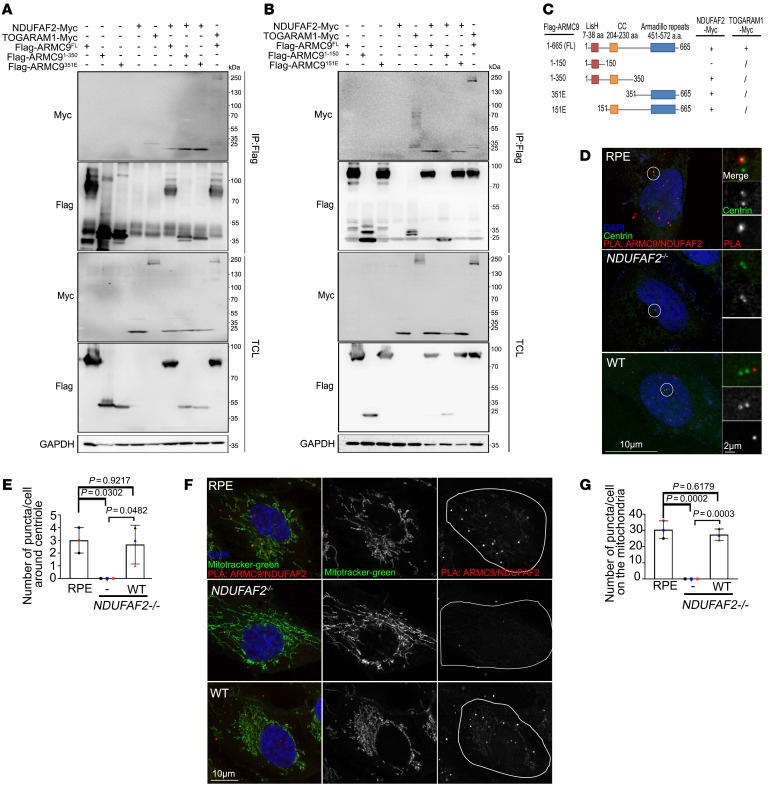
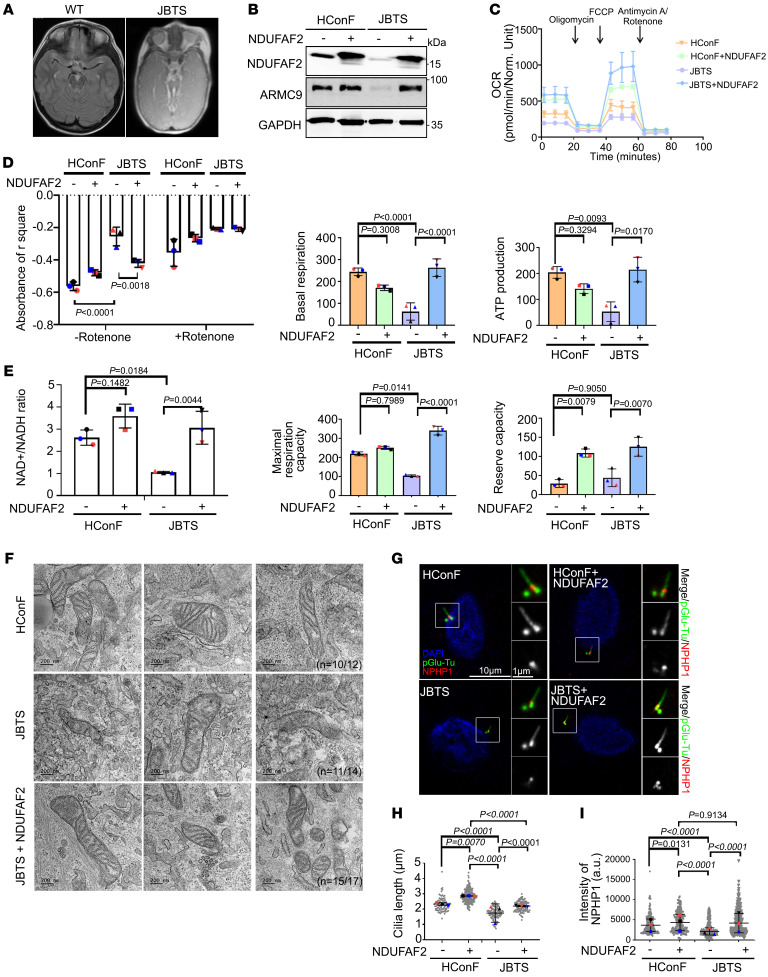
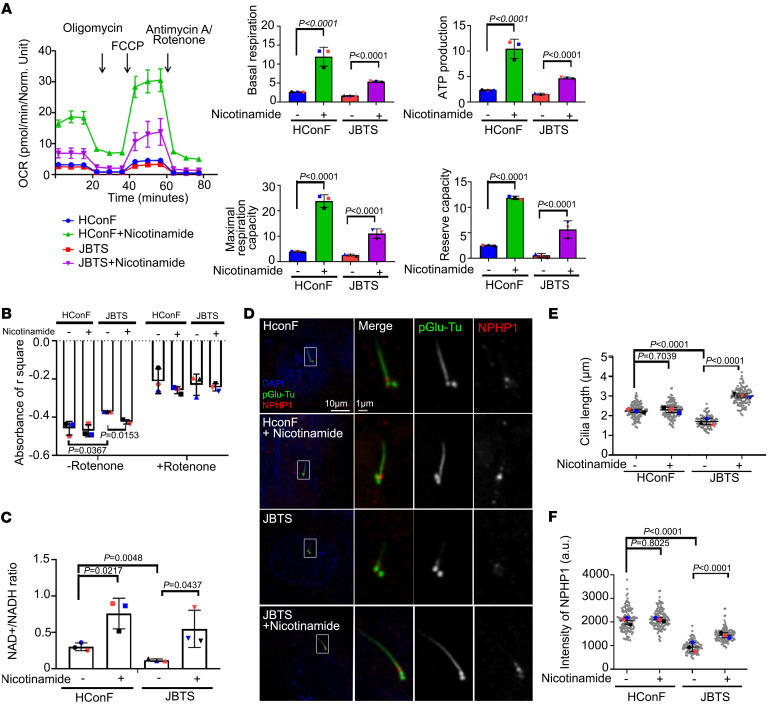
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
