

Distinct TORC1 signalling branches regulate Adc17 proteasome assembly chaperone expression
- PMID: 38949052
- PMCID: PMC11298713
- DOI: 10.1242/jcs.261892
Distinct TORC1 signalling branches regulate Adc17 proteasome assembly chaperone expression
Abstract
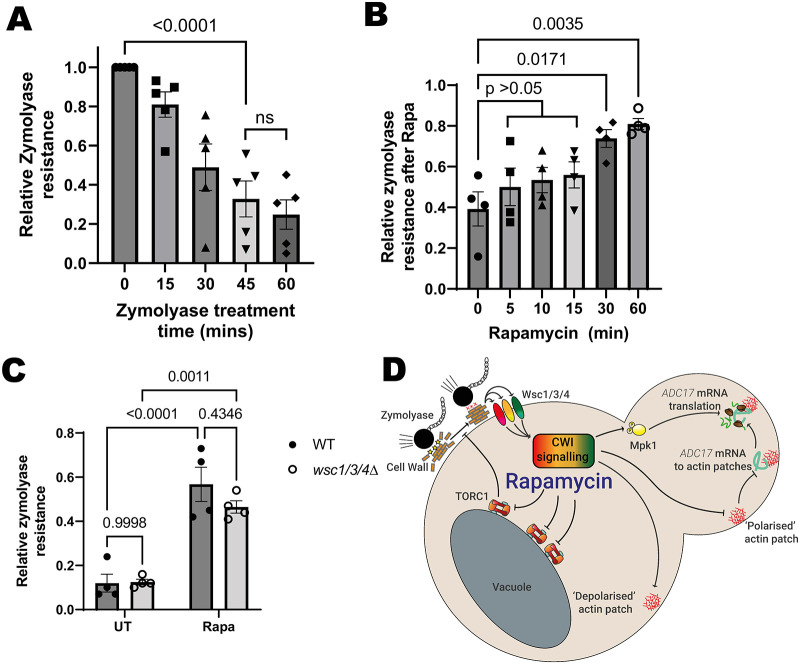
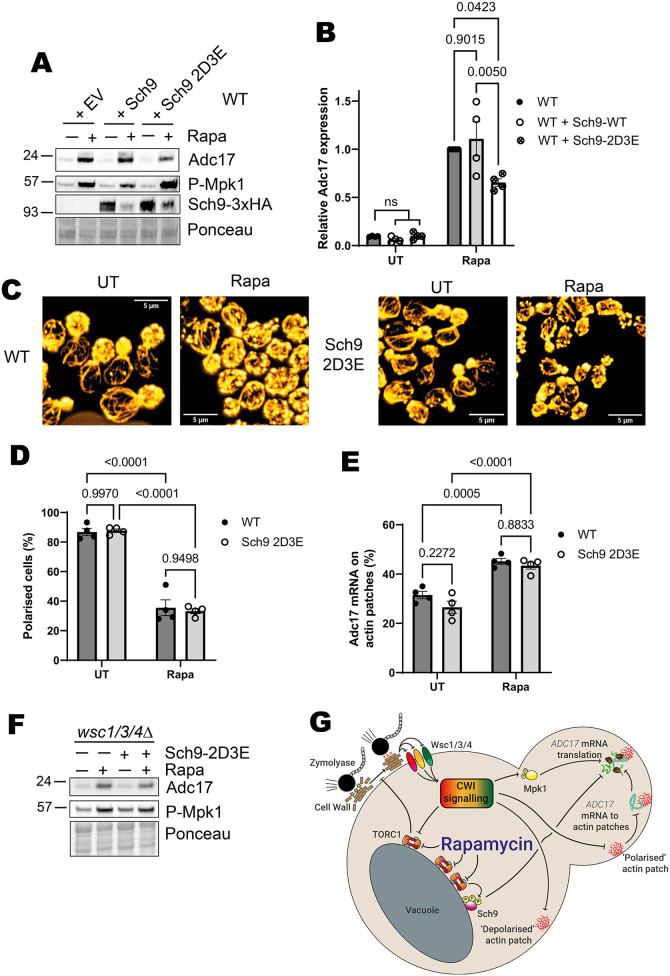
When stressed, cells need to adapt their proteome to maintain protein homeostasis. This requires increased proteasome assembly. Increased proteasome assembly is dependent on increased production of proteasome assembly chaperones. In Saccharomyces cerevisiae, inhibition of the growth-promoting kinase complex TORC1 causes increased proteasome assembly chaperone translation, including that of Adc17. This is dependent upon activation of the mitogen-activated protein kinase (MAPK) Mpk1 and relocalisation of assembly chaperone mRNA to patches of dense actin. We show here that TORC1 inhibition alters cell wall properties to induce these changes by activating the cell wall integrity pathway through the Wsc1, Wsc3 and Wsc4 sensor proteins. We demonstrate that, in isolation, these signals are insufficient to drive protein expression. We identify that the TORC1-activated S6 kinase Sch9 must be inhibited as well. This work expands our knowledge on the signalling pathways that regulate proteasome assembly chaperone production.
Keywords: Actin cytoskeleton; Mpk1 kinase; Proteasome assembly chaperone; Slt2 kinase; TORC1 signalling.
© 2024. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures





References
-
- Elorza, M. V., Rico, H. and Sentandreu, R. (1983). Calcofluor white alters the assembly of chitin fibrils in Saccharomyces cerevisiae and Candida albicans cells. J. Gen. Microbiol. 129, 1577-1582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
