

IFNγ-IL12 axis regulates intercellular crosstalk in metabolic dysfunction-associated steatotic liver disease
- PMID: 38951527
- PMCID: PMC11217362
- DOI: 10.1038/s41467-024-49633-y
IFNγ-IL12 axis regulates intercellular crosstalk in metabolic dysfunction-associated steatotic liver disease
Erratum in
-
Author Correction: IFNγ-IL12 axis regulates intercellular crosstalk in metabolic dysfunction-associated steatotic liver disease.Nat Commun. 2024 Jul 23;15(1):6195. doi: 10.1038/s41467-024-50607-3. Nat Commun. 2024. PMID: 39043691 Free PMC article. No abstract available.
Abstract
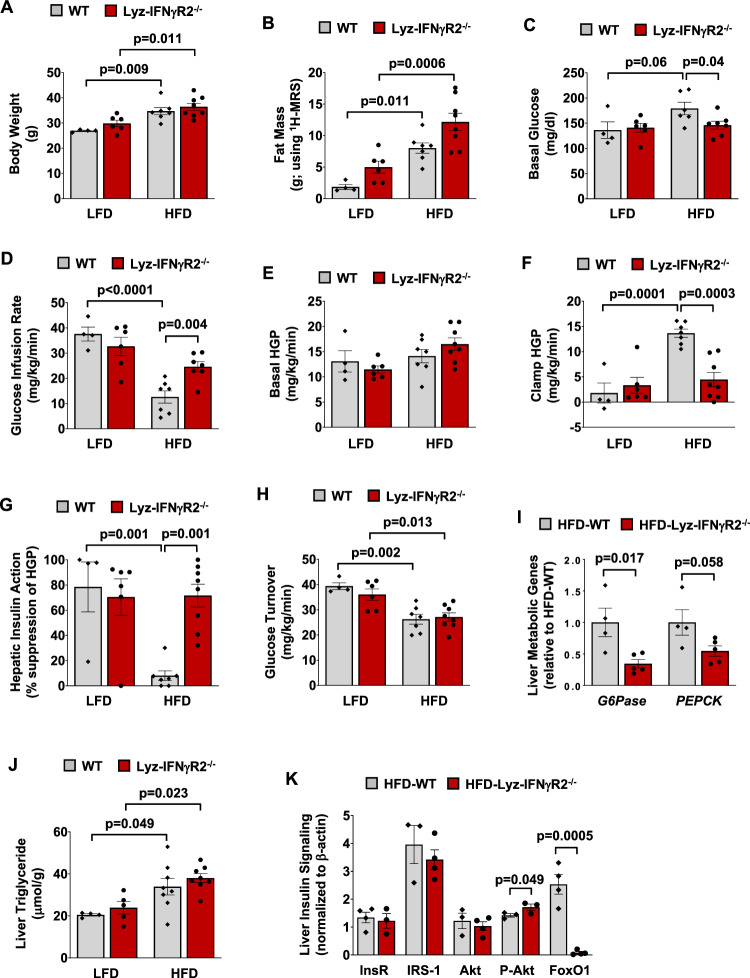
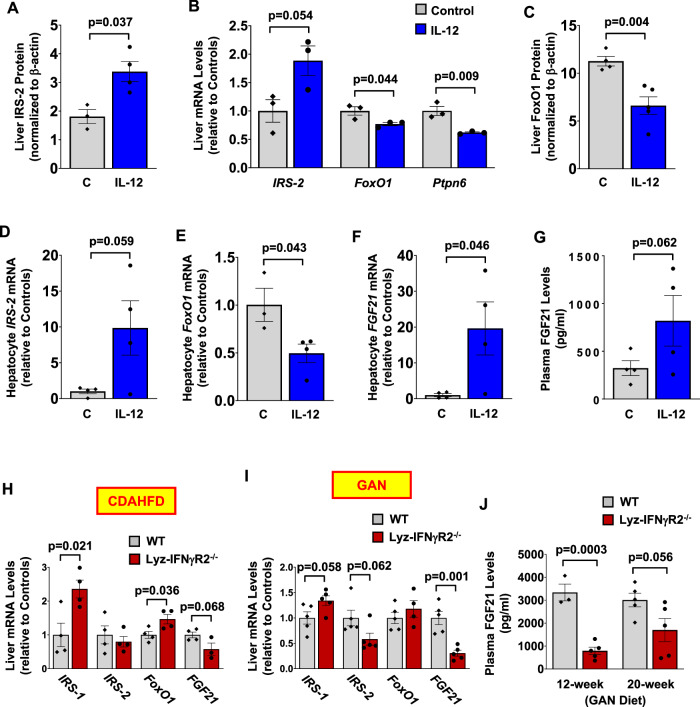
Obesity is a major cause of metabolic dysfunction-associated steatohepatitis (MASH) and is characterized by inflammation and insulin resistance. Interferon-γ (IFNγ) is a pro-inflammatory cytokine elevated in obesity and modulating macrophage functions. Here, we show that male mice with loss of IFNγ signaling in myeloid cells (Lyz-IFNγR2-/-) are protected from diet-induced insulin resistance despite fatty liver. Obesity-mediated liver inflammation is also attenuated with reduced interleukin (IL)-12, a cytokine primarily released by macrophages, and IL-12 treatment in vivo causes insulin resistance by impairing hepatic insulin signaling. Following MASH diets, Lyz-IFNγR2-/- mice are rescued from developing liver fibrosis, which is associated with reduced fibroblast growth factor (FGF) 21 levels. These results indicate critical roles for IFNγ signaling in macrophages and their release of IL-12 in modulating obesity-mediated insulin resistance and fatty liver progression to MASH. In this work, we identify the IFNγ-IL12 axis in regulating intercellular crosstalk in the liver and as potential therapeutic targets to treat MASH.
© 2024. The Author(s).
Conflict of interest statement
J.K.K. is on the scientific advisory board for Elevian Inc. and Imagine Pharma. D.L.G. is a consultant for The Jackson Laboratory. The remaining authors declare no competing interests.
Figures








References
MeSH terms
Substances
Grants and funding
- R01 DK133772/DK/NIDDK NIH HHS/United States
- UL1 TR001453/TR/NCATS NIH HHS/United States
- R01 DK116999/DK/NIDDK NIH HHS/United States
- R01DK116999/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- R01DK133772/U.S. Department of Health & Human Services | NIH | Center for Information Technology (Center for Information Technology, National Institutes of Health)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
