

Microscale geometrical modulation of PIEZO1 mediated mechanosensing through cytoskeletal redistribution
- PMID: 38951553
- PMCID: PMC11217425
- DOI: 10.1038/s41467-024-49833-6
Microscale geometrical modulation of PIEZO1 mediated mechanosensing through cytoskeletal redistribution
Abstract
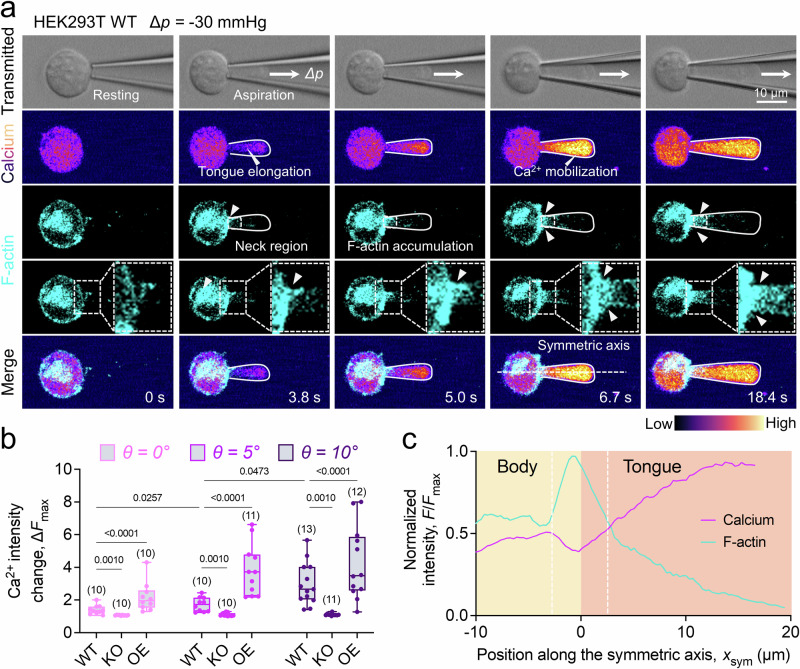
The microgeometry of the cellular microenvironment profoundly impacts cellular behaviors, yet the link between it and the ubiquitously expressed mechanosensitive ion channel PIEZO1 remains unclear. Herein, we describe a fluorescent micropipette aspiration assay that allows for simultaneous visualization of intracellular calcium dynamics and cytoskeletal architecture in real-time, under varied micropipette geometries. By integrating elastic shell finite element analysis with fluorescent lifetime imaging microscopy and employing PIEZO1-specific transgenic red blood cells and HEK cell lines, we demonstrate a direct correlation between the microscale geometry of aspiration and PIEZO1-mediated calcium signaling. We reveal that increased micropipette tip angles and physical constrictions lead to a significant reorganization of F-actin, accumulation at the aspirated cell neck, and subsequently amplify the tension stress at the dome of the cell to induce more PIEZO1's activity. Disruption of the F-actin network or inhibition of its mobility leads to a notable decline in PIEZO1 mediated calcium influx, underscoring its critical role in cellular mechanosensing amidst geometrical constraints.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
MeSH terms
Substances
Grants and funding
- DP200101970/Department of Education and Training | Australian Research Council (ARC)
- DP200101970/Department of Education and Training | Australian Research Council (ARC)
- APP2003904/Department of Health | National Health and Medical Research Council (NHMRC)
- APP2016165/Department of Health | National Health and Medical Research Council (NHMRC)
- APP1177374/Department of Health | National Health and Medical Research Council (NHMRC)
LinkOut - more resources
Full Text Sources
Miscellaneous
