

Does a Vertebrate Morphotype of Pallial Subdivisions Really Exist?
- PMID: 38952102
- PMCID: PMC11614313
- DOI: 10.1159/000537746
Does a Vertebrate Morphotype of Pallial Subdivisions Really Exist?
Abstract
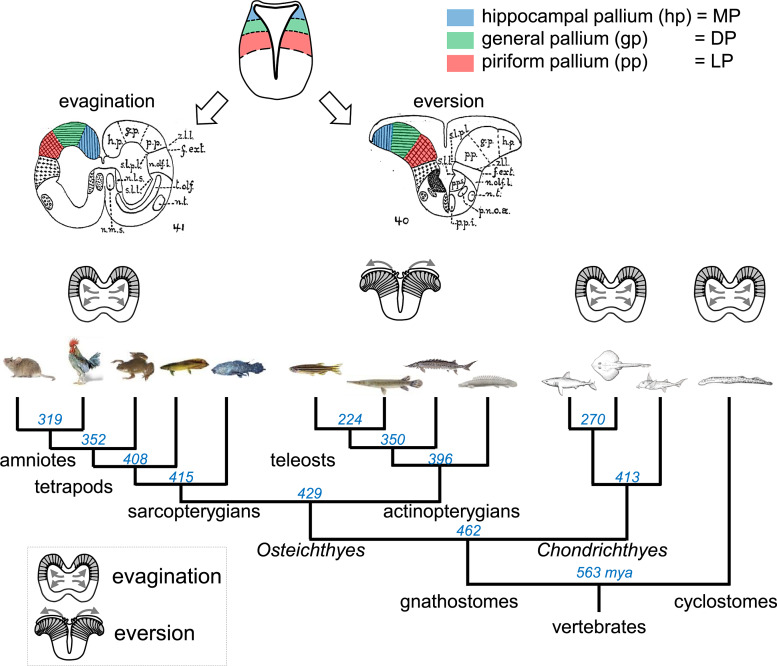
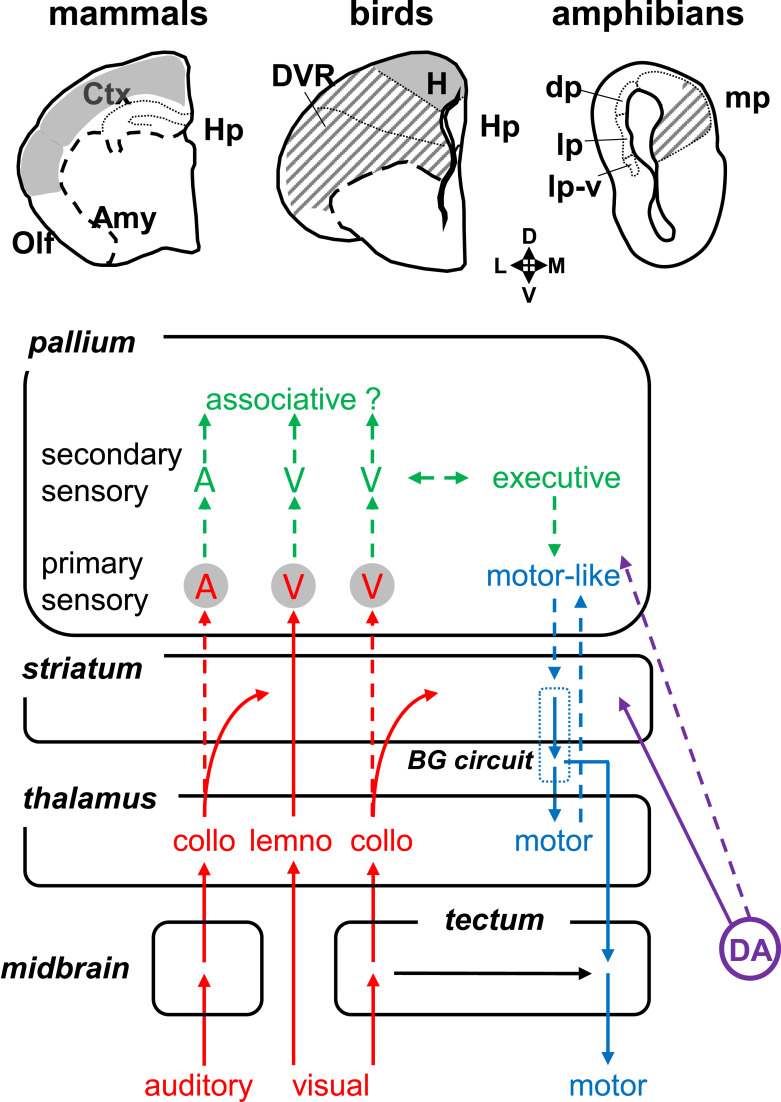
Background: Comparative neuroanatomists have long sought to determine which part of the pallium in nonmammals is homologous to the mammalian neocortex. A number of similar connectivity patterns across species have led to the idea that the basic organization of the vertebrate brain is relatively conserved; thus, efforts of the last decades have been focused on determining a vertebrate "morphotype" - a model comprising the characteristics believed to have been present in the last common ancestor of all vertebrates.
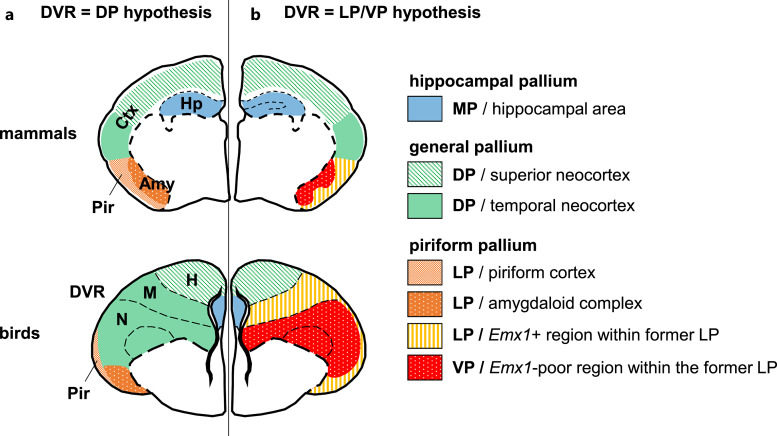
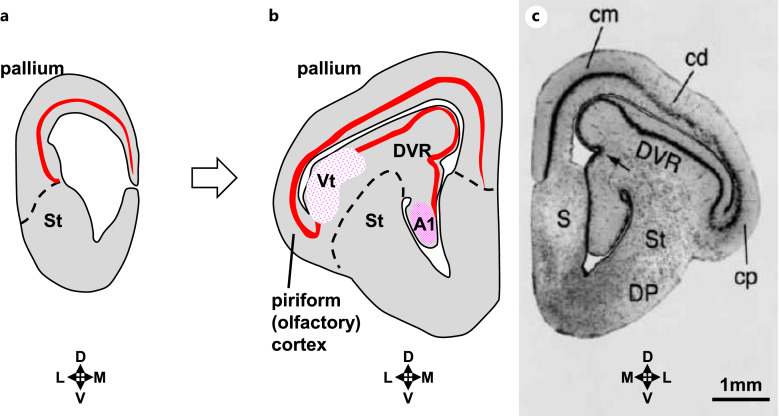
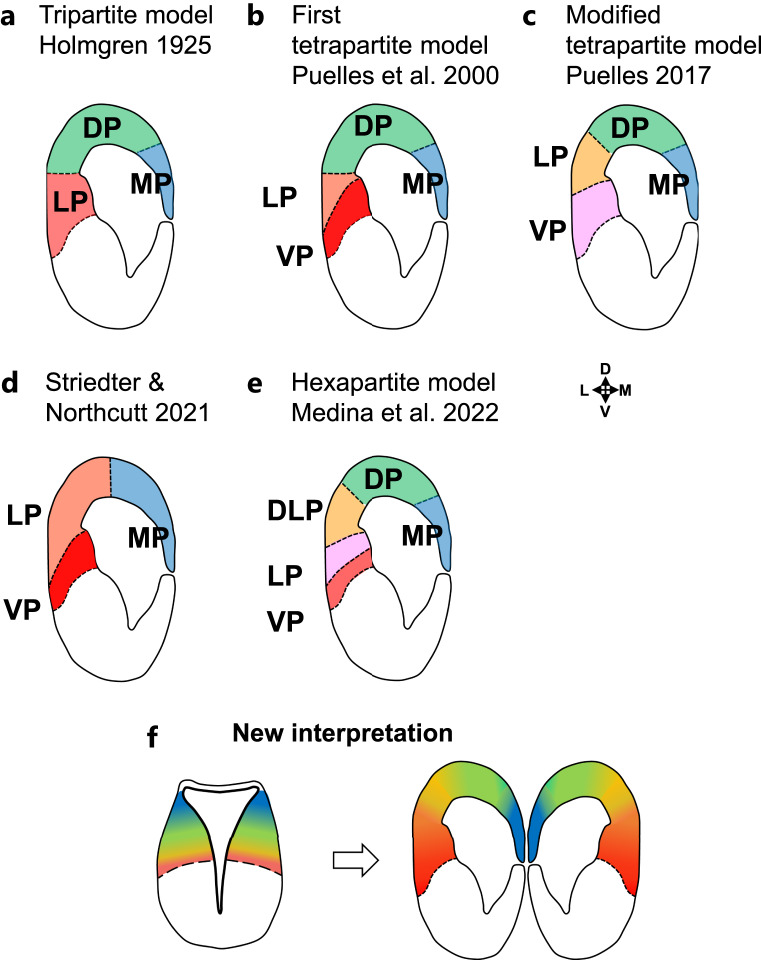
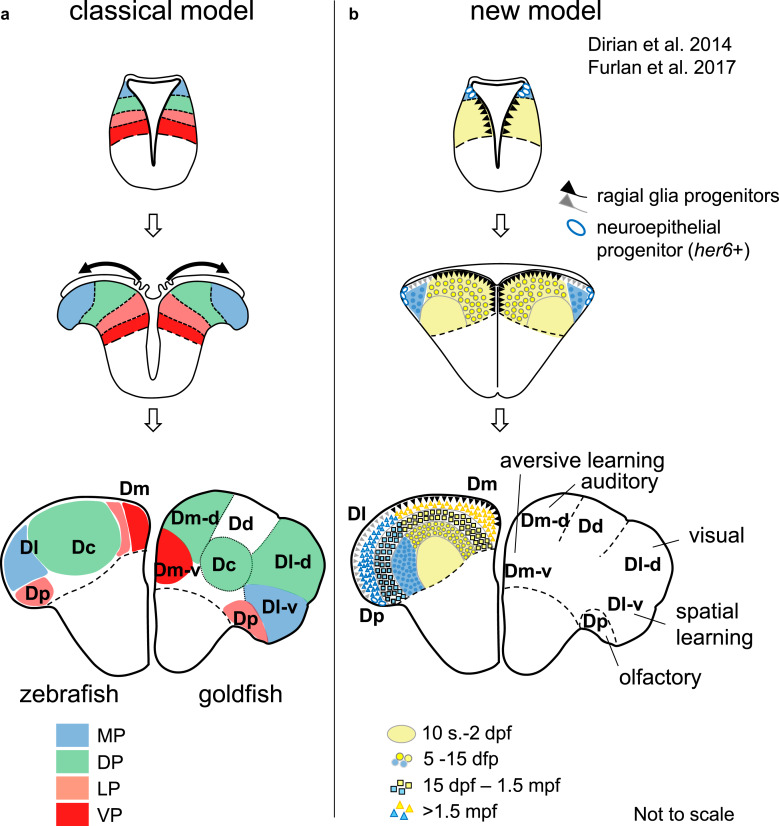
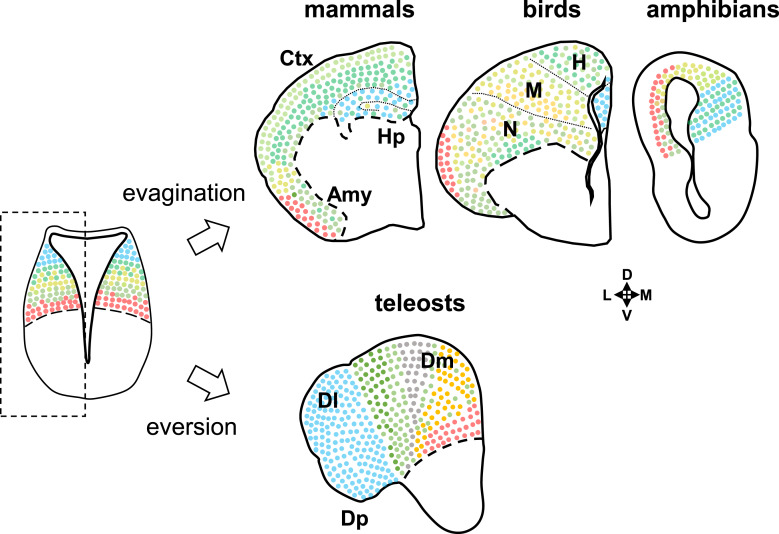
Summary: The endeavor to determine the vertebrate morphotype has been riddled with controversies due to the extensive morphological diversity of the pallium among vertebrate taxa. Nonetheless, most proposed scenarios of pallial homology are variants of a common theme where the vertebrate pallium is subdivided into subdivisions homologous to the hippocampus, neocortex, piriform cortex, and amygdala, in a one-to-one manner. We review the rationales of major propositions of pallial homology and identify the source of the discrepancies behind different hypotheses. We consider that a source of discrepancies is the prevailing assumption that there is a single "morphotype of the pallial subdivisions" throughout vertebrates. Instead, pallial subdivisions present in different taxa probably evolved independently in each lineage.
Key messages: We encounter discrepancies when we search for a single morphotype of subdivisions across vertebrates. These discrepancies can be resolved by considering that several subdivisions within the pallium were established after the divergence of the different lineages. The differences of pallial organization are especially remarkable between actinopterygians (including teleost fishes) and other vertebrates. Thus, the prevailing notion of a simple one-to-one homology between the mammalian and teleost pallia needs to be reconsidered.
Keywords: Comparative neuroanatomy; Convergent evolution; Homology; Pallium.
© 2024 The Author(s). Published by S. Karger AG, Basel.
Conflict of interest statement
The authors have no conflict of interests to declare.
Figures
Similar articles
-
Subdivisions of the adult zebrafish pallium based on molecular marker analysis.F1000Res. 2014 Dec 17;3:308. doi: 10.12688/f1000research.5595.2. eCollection 2014. F1000Res. 2014. PMID: 25713698 Free PMC article.
-
The Independent Evolution of Dorsal Pallia in Multiple Vertebrate Lineages.Brain Behav Evol. 2022;96(4-6):200-211. doi: 10.1159/000516563. Epub 2021 Jun 25. Brain Behav Evol. 2022. PMID: 34175847 Review.
-
Development and evolution of the pallium.Semin Cell Dev Biol. 2009 Aug;20(6):698-711. doi: 10.1016/j.semcdb.2009.04.008. Epub 2009 Apr 22. Semin Cell Dev Biol. 2009. PMID: 19393324 Review.
-
Spatial memory and hippocampal pallium through vertebrate evolution: insights from reptiles and teleost fish.Brain Res Bull. 2002 Feb-Mar 1;57(3-4):499-503. doi: 10.1016/s0361-9230(01)00682-7. Brain Res Bull. 2002. PMID: 11923018 Review.
-
Conservation and Diversification of Pallial Cell Types across Vertebrates: An Evo-Devo Perspective.Brain Behav Evol. 2023;98(4):210-228. doi: 10.1159/000531718. Epub 2023 Jun 28. Brain Behav Evol. 2023. PMID: 37379819 Review.
Cited by
-
The neural substrate of navigation using hydrostatic cues in goldfish.R Soc Open Sci. 2025 Feb 5;12(2):241869. doi: 10.1098/rsos.241869. eCollection 2025 Feb. R Soc Open Sci. 2025. PMID: 39911885 Free PMC article.
References
-
- Edinger L, Rand HW. The relations of comparative anatomy to comparative psychology. Comp Neurol Psychol. 1908;18(5):437–57.
-
- Ariëns Kappers CU. The phylogenesis of the paleo-cortex and archi-cortex compared with the evolution of the visual neo-cortex. Arch Neurol Psychiatry. 1909;4:161–73.
-
- Northcutt RG. The forebrain of gnathostomes: in search of a morphotype. Brain Behav Evol. 1995;46(4–5):275–318. - PubMed
-
- Holmgren N. Points of view concerning forebrain morphology in lower vertebrates. J Comp Neurol. 1922;34(5):391–459.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
