

Fusobacterium nucleatum subsp. nucleatum RadD binds Siglec-7 and inhibits NK cell-mediated cancer cell killing
- PMID: 38952680
- PMCID: PMC11215305
- DOI: 10.1016/j.isci.2024.110157
Fusobacterium nucleatum subsp. nucleatum RadD binds Siglec-7 and inhibits NK cell-mediated cancer cell killing
Abstract
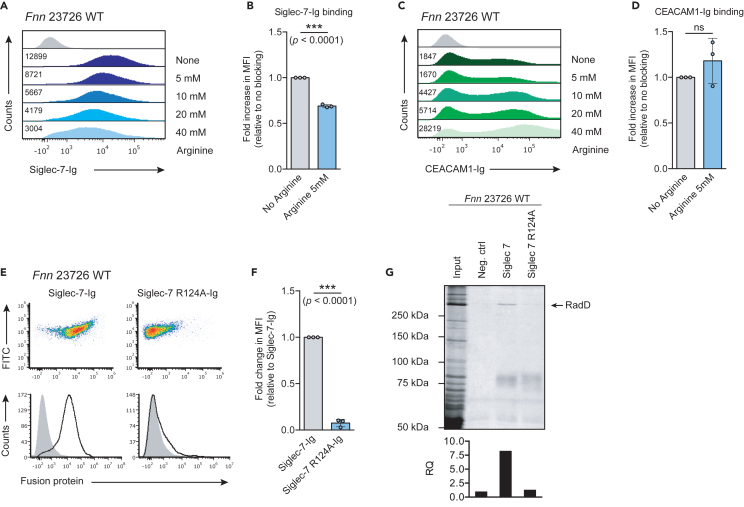
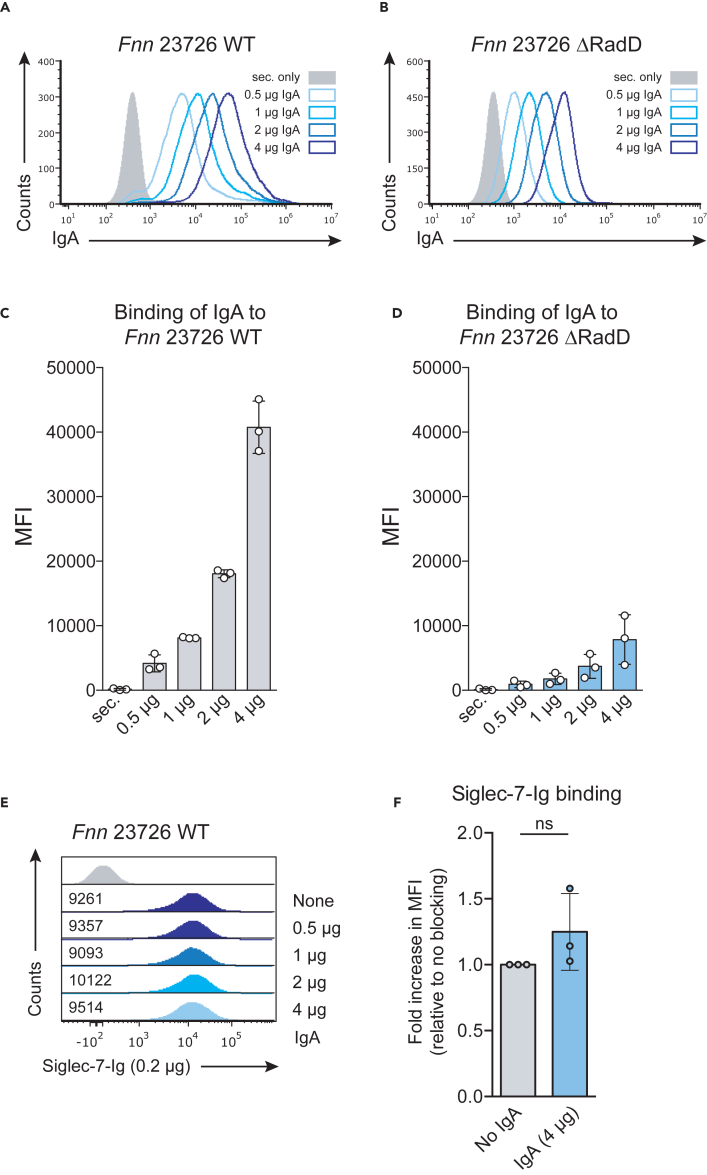
Fusobacterium nucleatum is an oral commensal bacterium that can colonize extraoral tumor entities, such as colorectal cancer and breast cancer. Recent studies revealed its ability to modulate the immune response in the tumor microenvironment (TME), promoting cancer progression and metastasis. Importantly, F. nucleatum subsp. animalis was shown to bind to Siglec-7 via lipopolysaccharides, leading to a pro-inflammatory profile in human monocyte-derived dendritic cells. In this study, we show that F. nucleatum subsp. nucleatum RadD binds to Siglec-7 on NK cells, thereby inhibiting NK cell-mediated cancer cell killing. We demonstrate that this binding is dependent on arginine residue R124 in Siglec-7. Finally, we determine that this binding is independent of the known interaction of RadD with IgA. Taken together, our findings elucidate the targeting of Siglec-7 by F. nucleatum subsp. nucleatum RadD as a means to modulate the NK cell response and potentially promoting immune evasion and tumor progression.
Keywords: cancer; immunology; microbiology; molecular biology.
© 2024 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Galeano Niño J.L., Wu H., LaCourse K.D., Kempchinsky A.G., Baryiames A., Barber B., Futran N., Houlton J., Sather C., Sicinska E., et al. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature. 2022;611:810–817. doi: 10.1038/s41586-022-05435-0. - DOI - PMC - PubMed
-
- Yamamura K., Baba Y., Nakagawa S., Mima K., Miyake K., Nakamura K., Sawayama H., Kinoshita K., Ishimoto T., Iwatsuki M., et al. Human Microbiome Fusobacterium Nucleatum in Esophageal Cancer Tissue Is Associated with Prognosis. Clin. Cancer Res. 2016;22:5574–5581. doi: 10.1158/1078-0432.ccr-16-1786. - DOI - PubMed
-
- Mitsuhashi K., Nosho K., Sukawa Y., Matsunaga Y., Ito M., Kurihara H., Kanno S., Igarashi H., Naito T., Adachi Y., et al. Association of Fusobacterium species in pancreatic cancer tissues with molecular features and prognosis. Oncotarget. 2015;6:7209–7220. doi: 10.18632/oncotarget.3109. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
