

Inflammation primes the murine kidney for recovery by activating AZIN1 adenosine-to-inosine editing
- PMID: 38954486
- PMCID: PMC11364396
- DOI: 10.1172/JCI180117
Inflammation primes the murine kidney for recovery by activating AZIN1 adenosine-to-inosine editing
Abstract
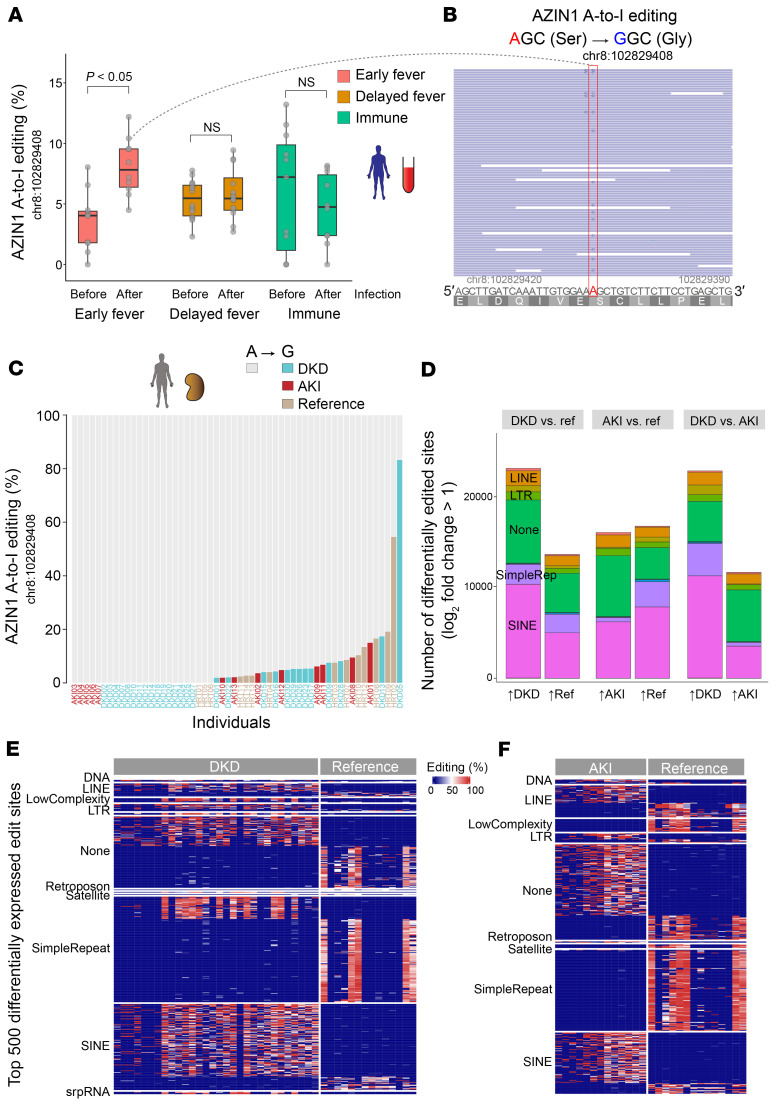
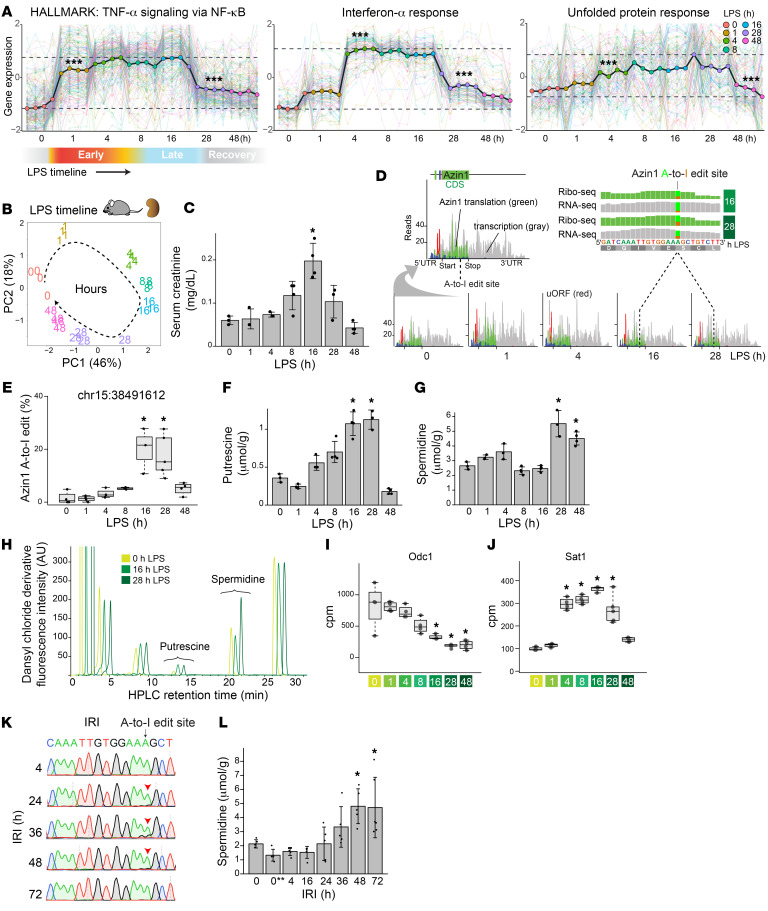
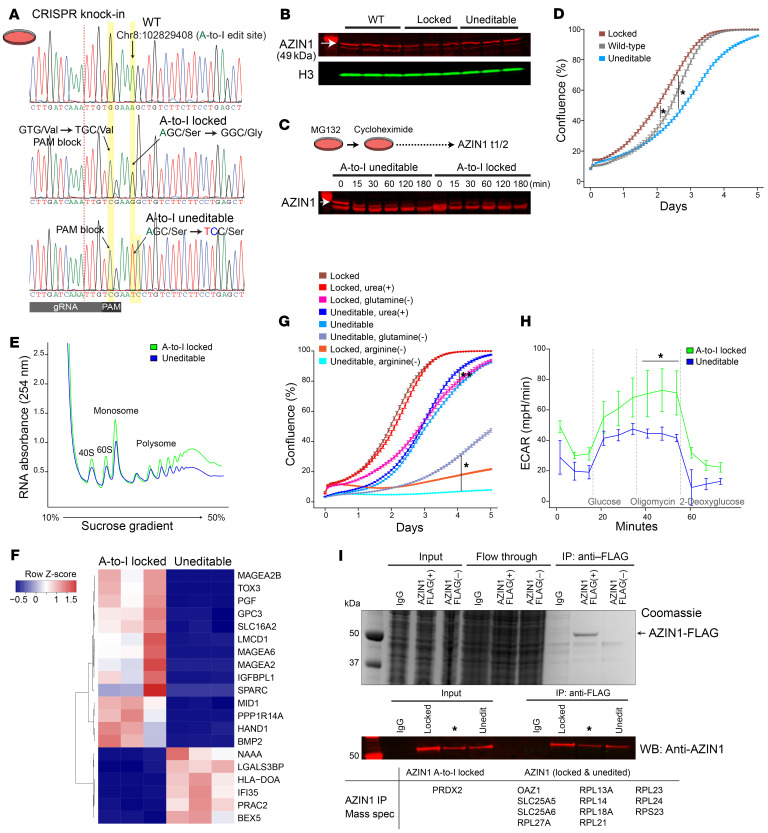
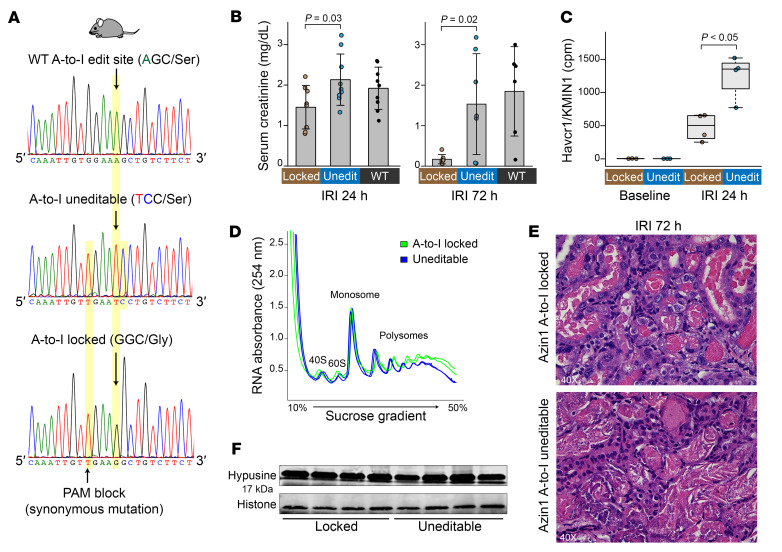
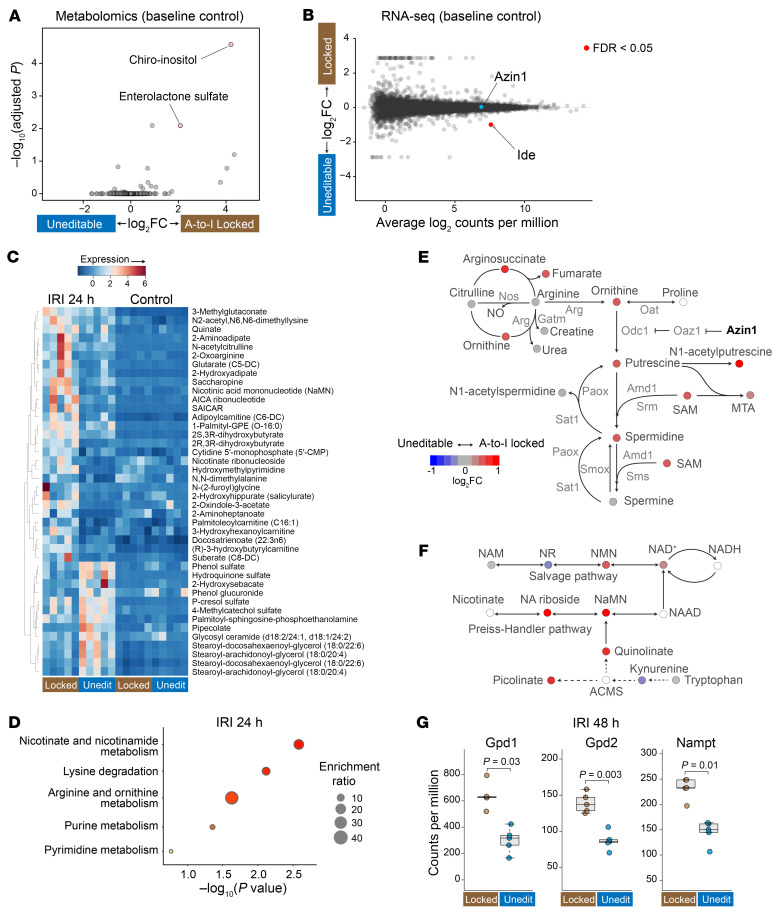
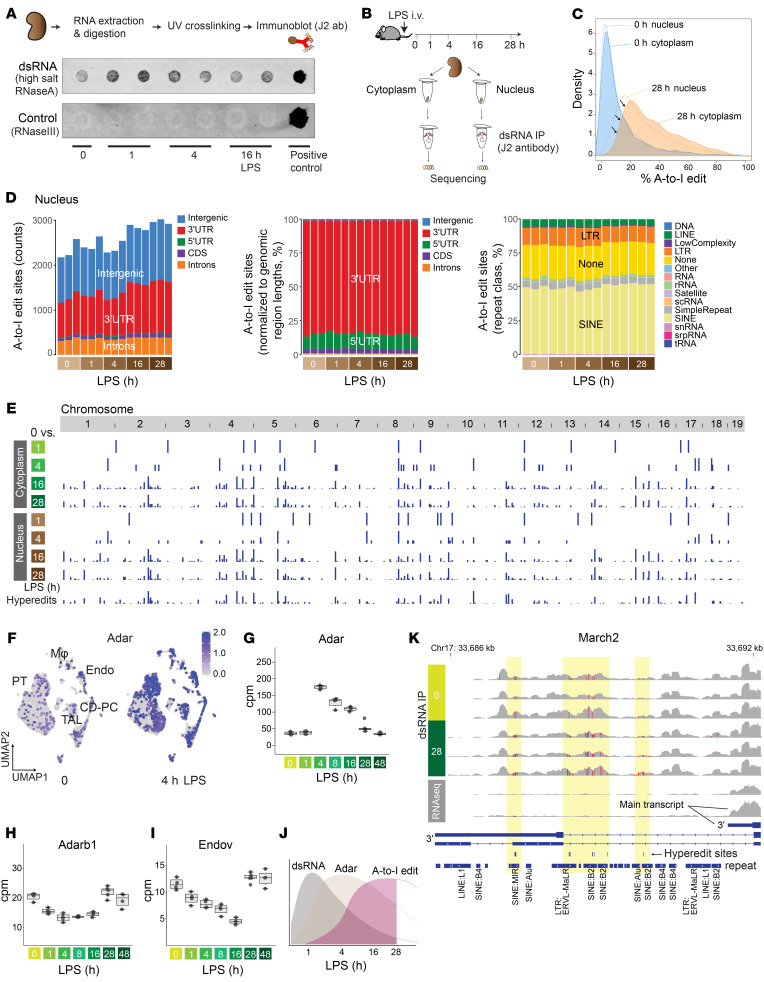
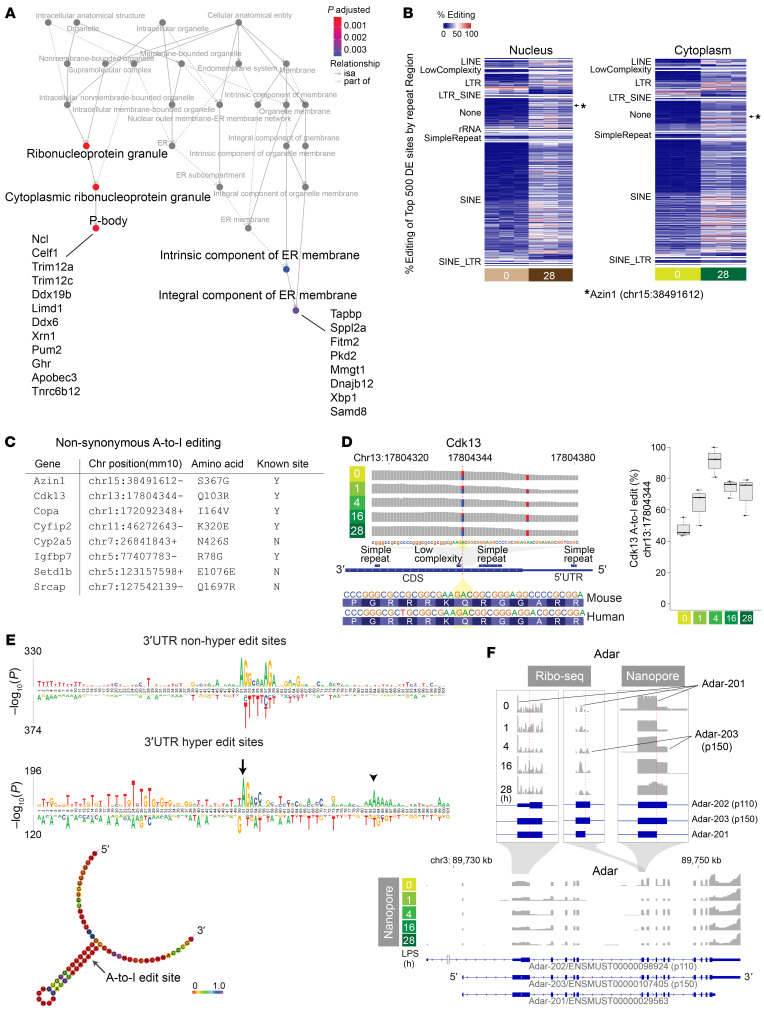
The progression of kidney disease varies among individuals, but a general methodology to quantify disease timelines is lacking. Particularly challenging is the task of determining the potential for recovery from acute kidney injury following various insults. Here, we report that quantitation of post-transcriptional adenosine-to-inosine (A-to-I) RNA editing offers a distinct genome-wide signature, enabling the delineation of disease trajectories in the kidney. A well-defined murine model of endotoxemia permitted the identification of the origin and extent of A-to-I editing, along with temporally discrete signatures of double-stranded RNA stress and adenosine deaminase isoform switching. We found that A-to-I editing of antizyme inhibitor 1 (AZIN1), a positive regulator of polyamine biosynthesis, serves as a particularly useful temporal landmark during endotoxemia. Our data indicate that AZIN1 A-to-I editing, triggered by preceding inflammation, primes the kidney and activates endogenous recovery mechanisms. By comparing genetically modified human cell lines and mice locked in either A-to-I-edited or uneditable states, we uncovered that AZIN1 A-to-I editing not only enhances polyamine biosynthesis but also engages glycolysis and nicotinamide biosynthesis to drive the recovery phenotype. Our findings implicate that quantifying AZIN1 A-to-I editing could potentially identify individuals who have transitioned to an endogenous recovery phase. This phase would reflect their past inflammation and indicate their potential for future recovery.
Keywords: Bioinformatics; Cell stress; Nephrology; Polyamines.
Conflict of interest statement
Figures
Update of
-
Inflammation primes the kidney for recovery by activating AZIN1 A-to-I editing.bioRxiv [Preprint]. 2023 Nov 9:2023.11.09.566426. doi: 10.1101/2023.11.09.566426. bioRxiv. 2023. Update in: J Clin Invest. 2024 Sep 3;134(17):e180117. doi: 10.1172/JCI180117. PMID: 37986799 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
