

Intrinsic signaling pathways modulate targeted protein degradation
- PMID: 38956052
- PMCID: PMC11220168
- DOI: 10.1038/s41467-024-49519-z
Intrinsic signaling pathways modulate targeted protein degradation
Abstract
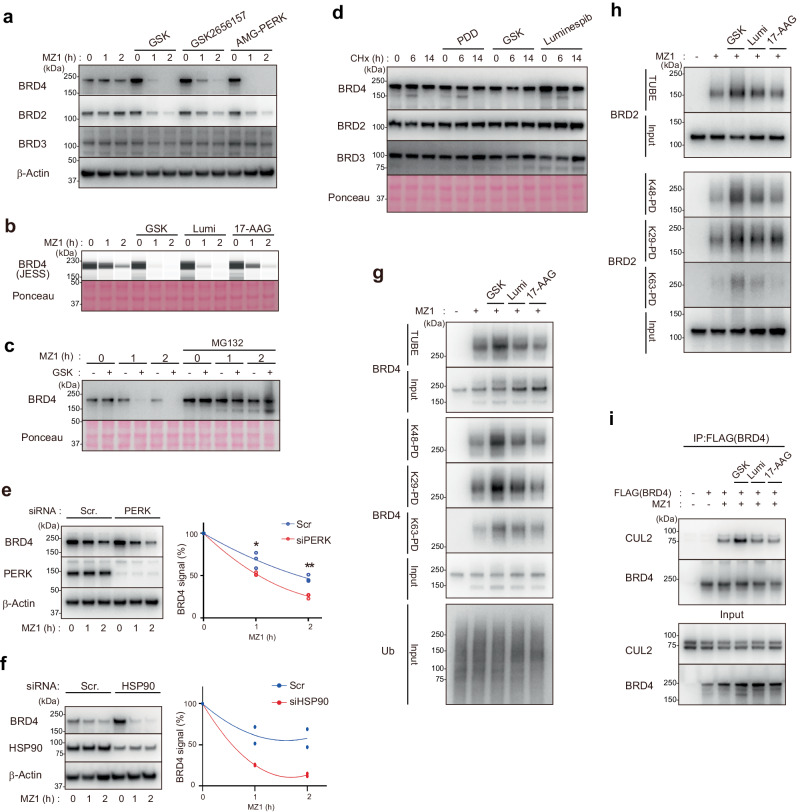
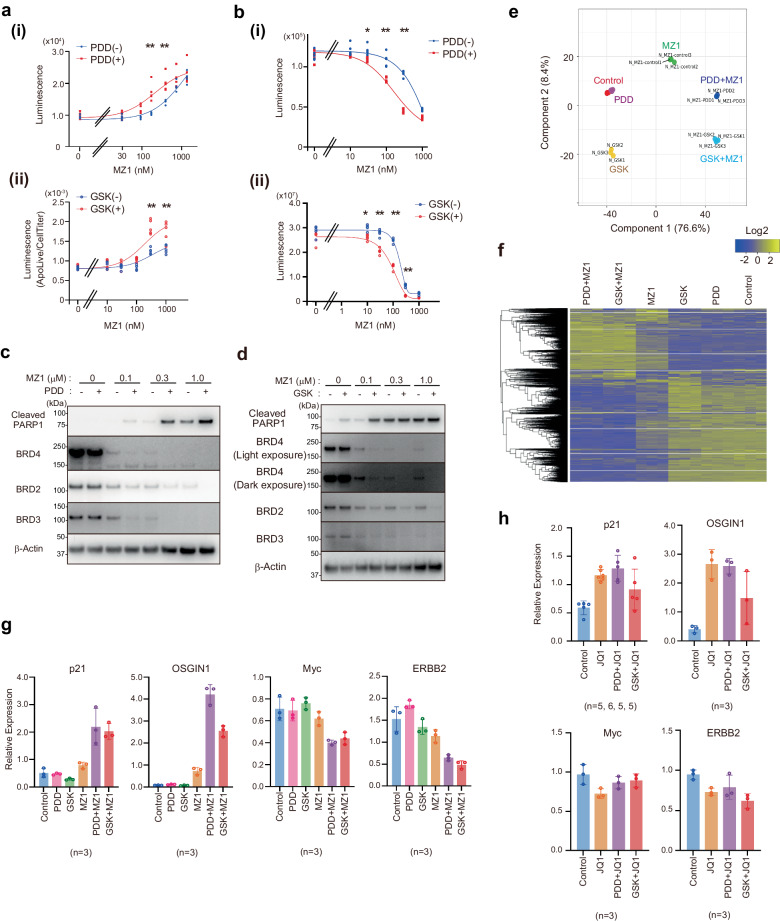
Targeted protein degradation is a groundbreaking modality in drug discovery; however, the regulatory mechanisms are still not fully understood. Here, we identify cellular signaling pathways that modulate the targeted degradation of the anticancer target BRD4 and related neosubstrates BRD2/3 and CDK9 induced by CRL2VHL- or CRL4CRBN -based PROTACs. The chemicals identified as degradation enhancers include inhibitors of cellular signaling pathways such as poly-ADP ribosylation (PARG inhibitor PDD00017273), unfolded protein response (PERK inhibitor GSK2606414), and protein stabilization (HSP90 inhibitor luminespib). Mechanistically, PARG inhibition promotes TRIP12-mediated K29/K48-linked branched ubiquitylation of BRD4 by facilitating chromatin dissociation of BRD4 and formation of the BRD4-PROTAC-CRL2VHL ternary complex; by contrast, HSP90 inhibition promotes BRD4 degradation after the ubiquitylation step. Consequently, these signal inhibitors sensitize cells to the PROTAC-induced apoptosis. These results suggest that various cell-intrinsic signaling pathways spontaneously counteract chemically induced target degradation at multiple steps, which could be liberated by specific inhibitors.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
