

A rapid aureochrome opto-switch enables diatom acclimation to dynamic light
- PMID: 38956103
- PMCID: PMC11219949
- DOI: 10.1038/s41467-024-49991-7
A rapid aureochrome opto-switch enables diatom acclimation to dynamic light
Abstract
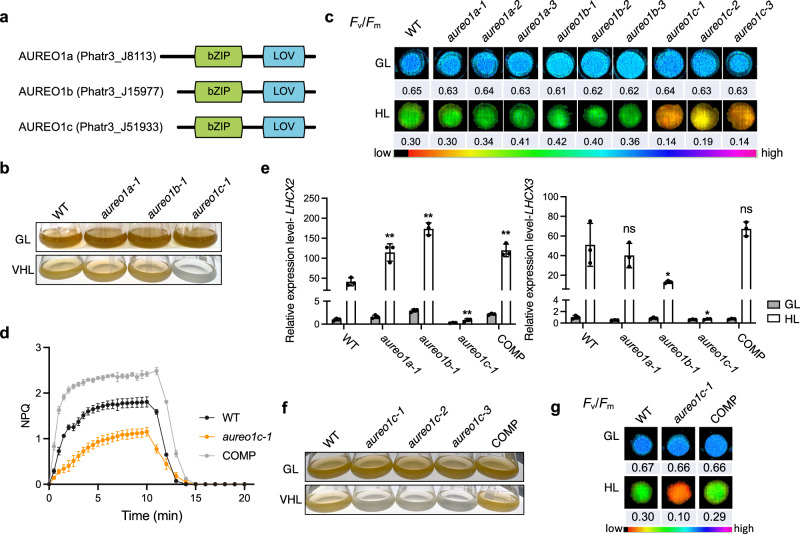
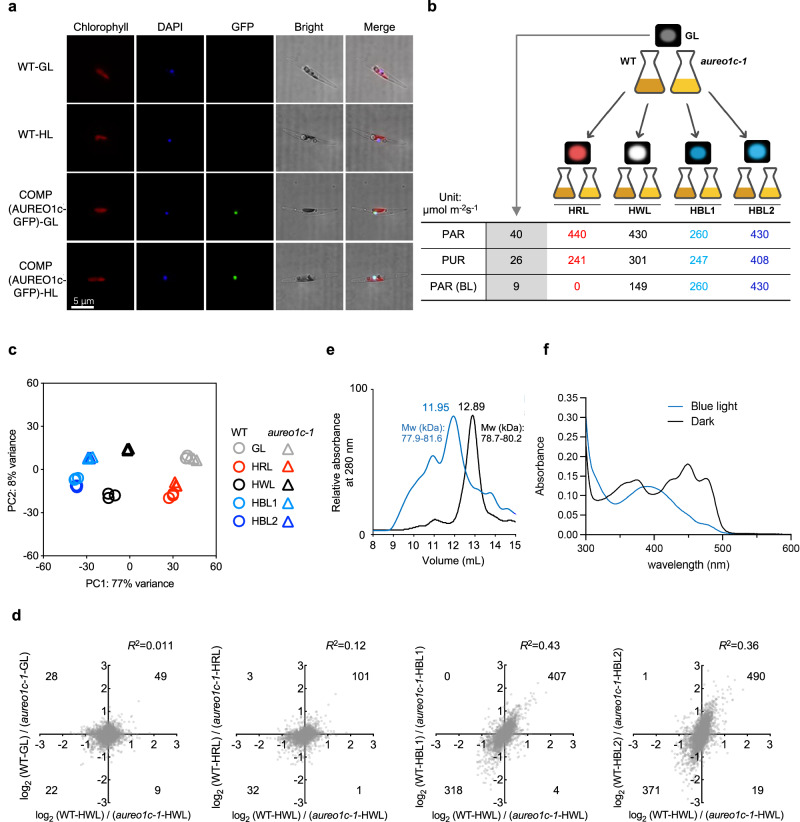
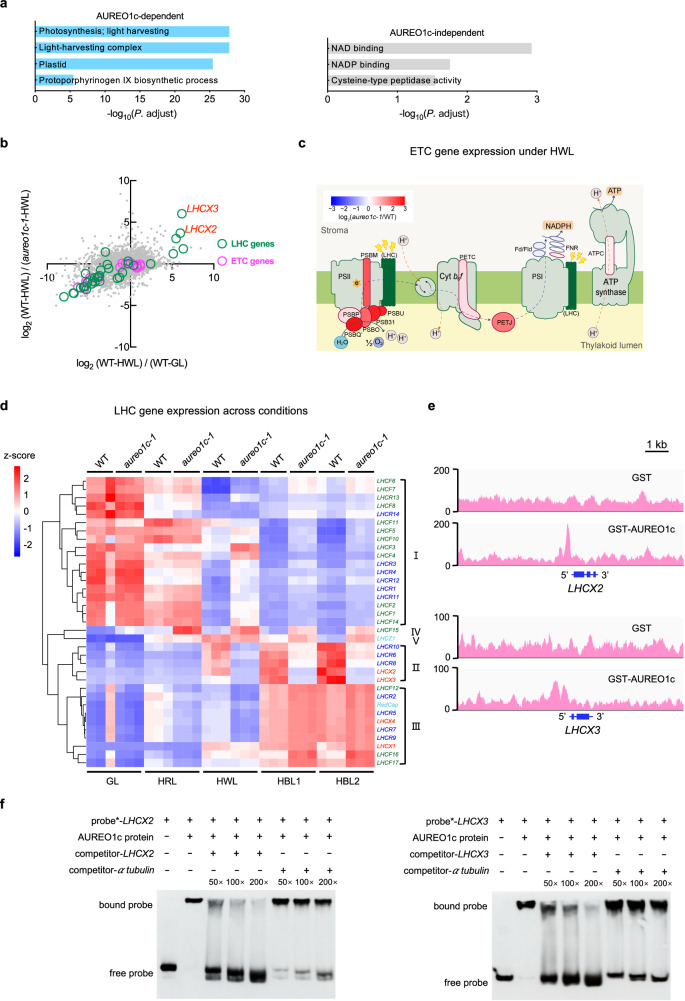
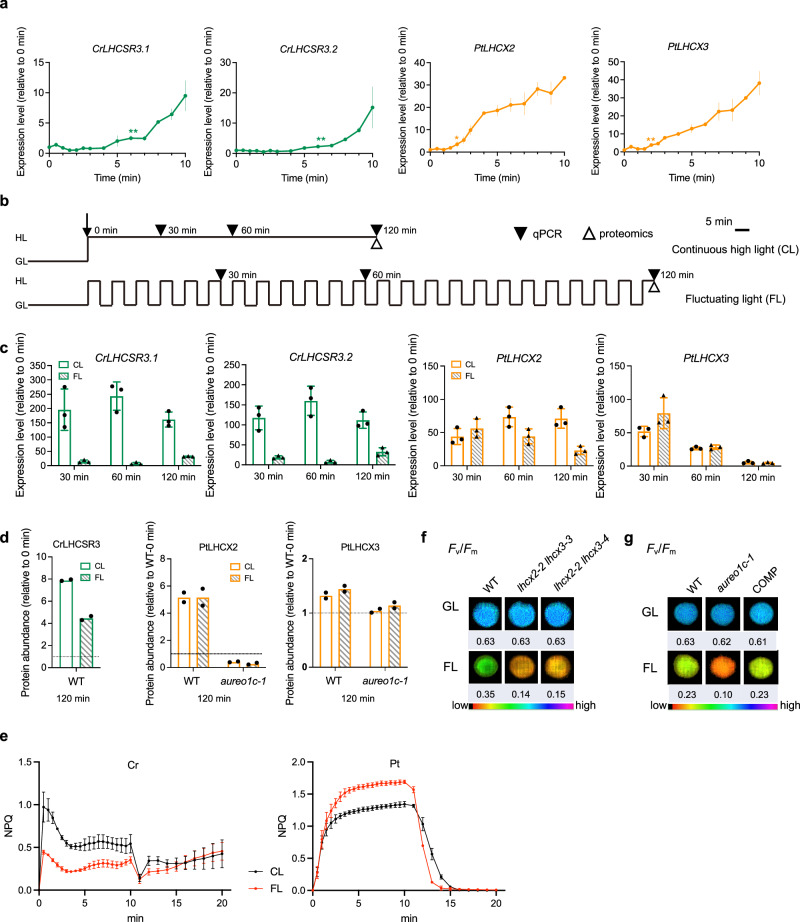
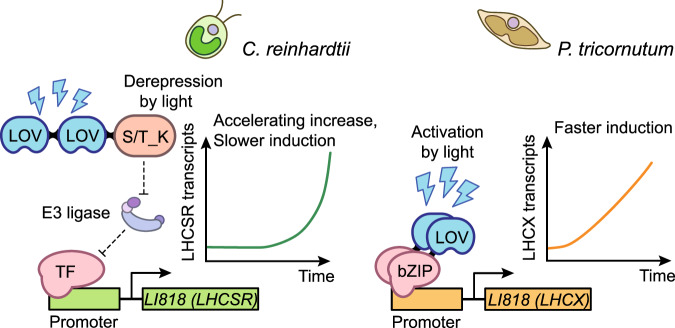
Diatoms often outnumber other eukaryotic algae in the oceans, especially in coastal environments characterized by frequent fluctuations in light intensity. The identities and operational mechanisms of regulatory factors governing diatom acclimation to high light stress remain largely elusive. Here, we identified the AUREO1c protein from the coastal diatom Phaeodactylum tricornutum as a crucial regulator of non-photochemical quenching (NPQ), a photoprotective mechanism that dissipates excess energy as heat. AUREO1c detects light stress using a light-oxygen-voltage (LOV) domain and directly activates the expression of target genes, including LI818 genes that encode NPQ effector proteins, via its bZIP DNA-binding domain. In comparison to a kinase-mediated pathway reported in the freshwater green alga Chlamydomonas reinhardtii, the AUREO1c pathway exhibits a faster response and enables accumulation of LI818 transcript and protein levels to comparable degrees between continuous high-light and fluctuating-light treatments. We propose that the AUREO1c-LI818 pathway contributes to the resilience of diatoms under dynamic light conditions.
© 2024. The Author(s).
Conflict of interest statement
X.L., H.Z., K.G., X.X., and T.C. have submitted a Chinese patent application for this work presented in the manuscript. The application number is 202410675502.8. The other authors declare that they have no competing interests.
Figures
Similar articles
-
Multisignal control of expression of the LHCX protein family in the marine diatom Phaeodactylum tricornutum.J Exp Bot. 2016 Jun;67(13):3939-51. doi: 10.1093/jxb/erw198. Epub 2016 May 25. J Exp Bot. 2016. PMID: 27225826 Free PMC article.
-
PsbS contributes to photoprotection in Chlamydomonas reinhardtii independently of energy dissipation.Biochim Biophys Acta Bioenerg. 2020 Jun 1;1861(5-6):148183. doi: 10.1016/j.bbabio.2020.148183. Epub 2020 Mar 12. Biochim Biophys Acta Bioenerg. 2020. PMID: 32173384
-
Chlamydomonas reinhardtii PsbS Protein Is Functional and Accumulates Rapidly and Transiently under High Light.Plant Physiol. 2016 Aug;171(4):2717-30. doi: 10.1104/pp.16.00572. Epub 2016 Jun 21. Plant Physiol. 2016. PMID: 27329221 Free PMC article.
-
Non-Photochemical Quenching: From Light Perception to Photoprotective Gene Expression.Int J Mol Sci. 2022 Jan 8;23(2):687. doi: 10.3390/ijms23020687. Int J Mol Sci. 2022. PMID: 35054872 Free PMC article. Review.
-
An update on aureochromes: Phylogeny - mechanism - function.J Plant Physiol. 2017 Oct;217:20-26. doi: 10.1016/j.jplph.2017.06.010. Epub 2017 Jul 11. J Plant Physiol. 2017. PMID: 28797596 Review.
Cited by
-
Regulation of Microalgal Photosynthetic Electron Transfer.Plants (Basel). 2024 Jul 29;13(15):2103. doi: 10.3390/plants13152103. Plants (Basel). 2024. PMID: 39124221 Free PMC article. Review.
-
Night-break treatment with blue and green lights regulates erect thallus formation in the brown alga Petalonia fascia (KU-1293).Front Plant Sci. 2025 Jan 15;15:1500947. doi: 10.3389/fpls.2024.1500947. eCollection 2024. Front Plant Sci. 2025. PMID: 39881733 Free PMC article.
-
Uridine diphosphate glucose confers oxidative stress tolerance in microalgae.BMC Plant Biol. 2025 May 1;25(1):566. doi: 10.1186/s12870-025-06587-7. BMC Plant Biol. 2025. PMID: 40307684 Free PMC article.
References
-
- Katz ME, Finkel ZV, Grzebyk D, Knoll AH, Falkowski PG. Evolutionary trajectories and biogeochemical impacts of marine eukaryotic phytoplankton. Annu. Rev. Ecol. Evol. Syst. 2004;35:523–556. doi: 10.1146/annurev.ecolsys.35.112202.130137. - DOI
-
- Seth K, et al. Bioprospecting of fucoxanthin from diatoms—challenges and perspectives. Algal Res. 2021;60:102475. doi: 10.1016/j.algal.2021.102475. - DOI
-
- Wang, W. et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science363, 10.1126/science.aav0365 (2019). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
