

Environmental circadian disruption re-writes liver circadian proteomes
- PMID: 38956413
- PMCID: PMC11220080
- DOI: 10.1038/s41467-024-49852-3
Environmental circadian disruption re-writes liver circadian proteomes
Abstract
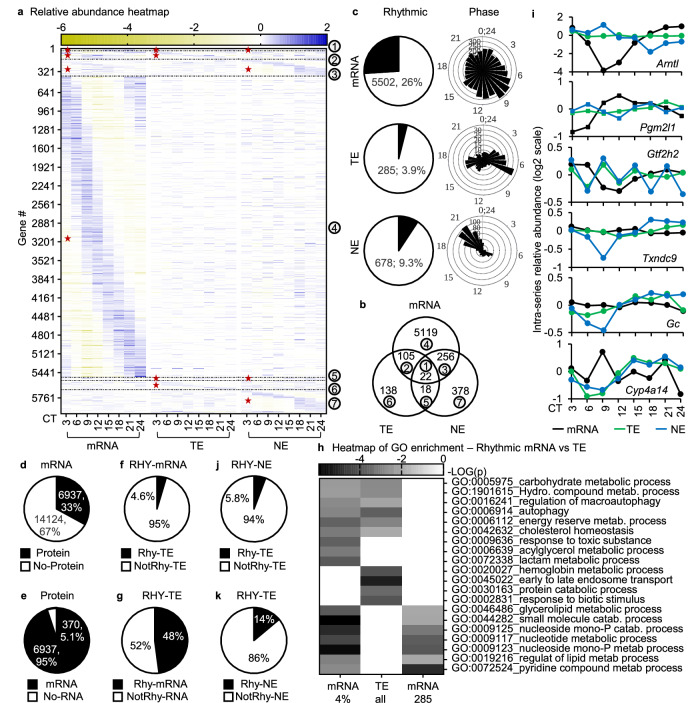
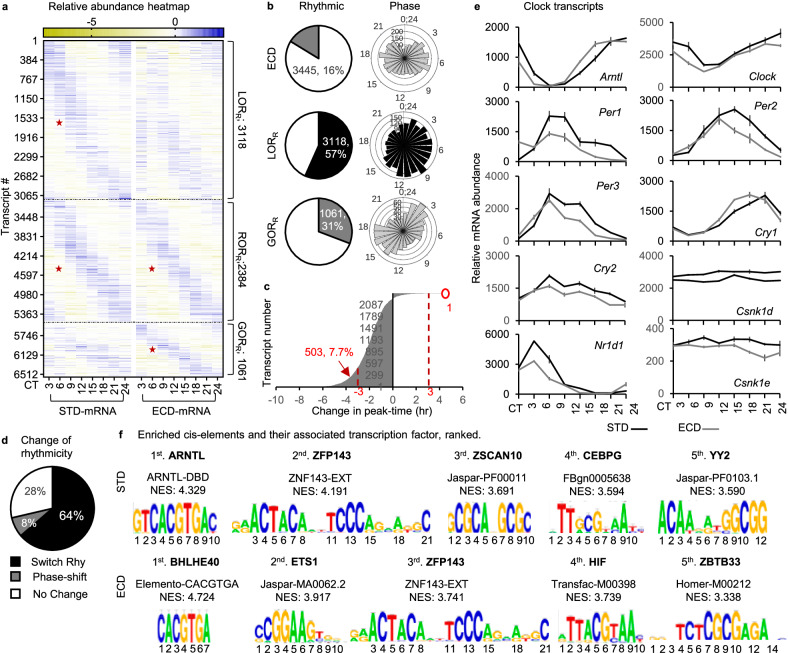
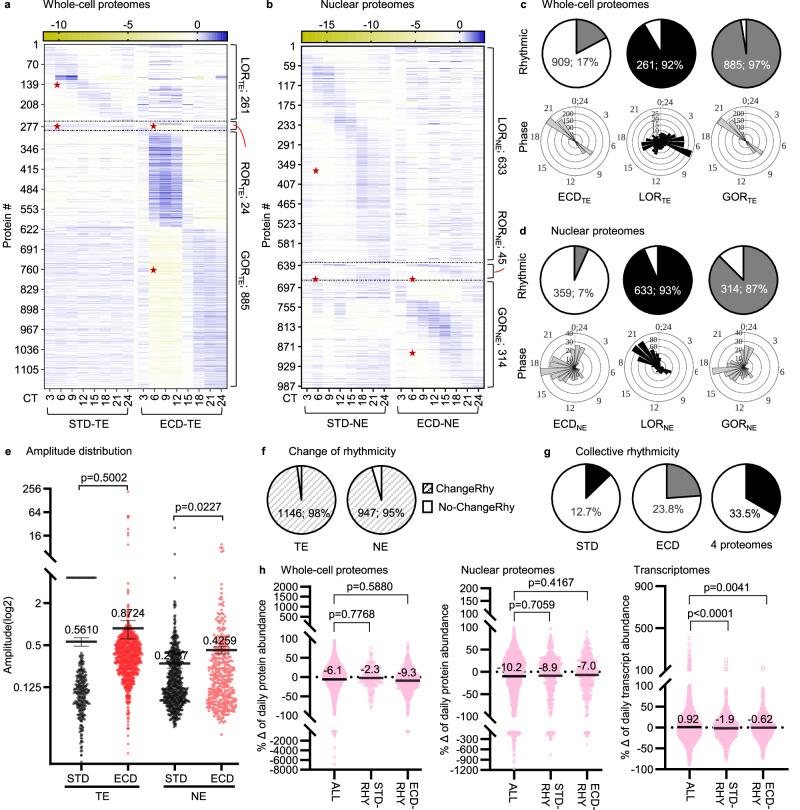
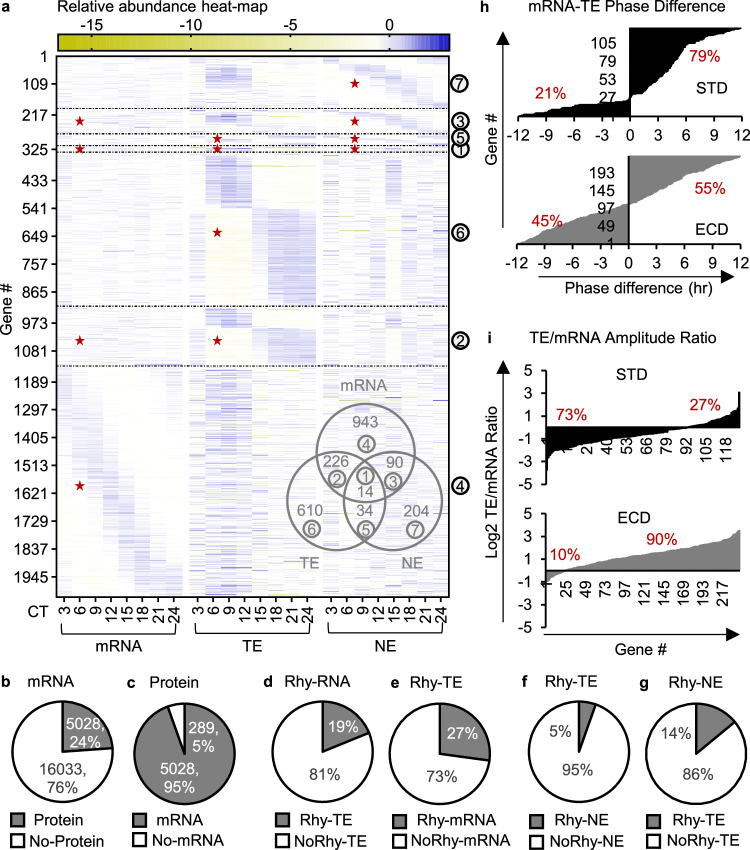
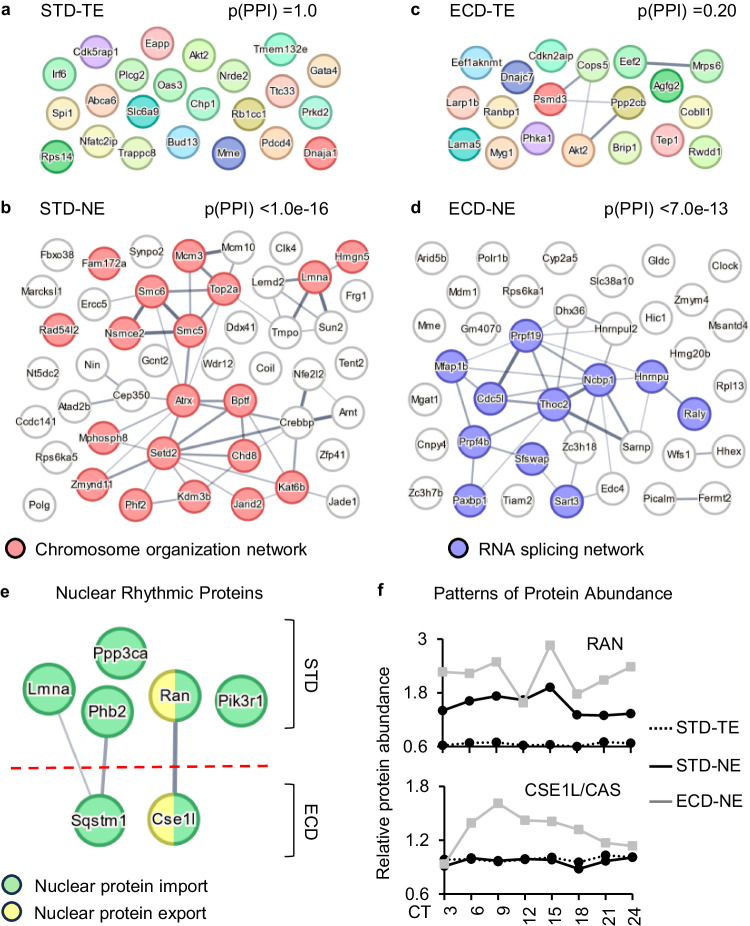
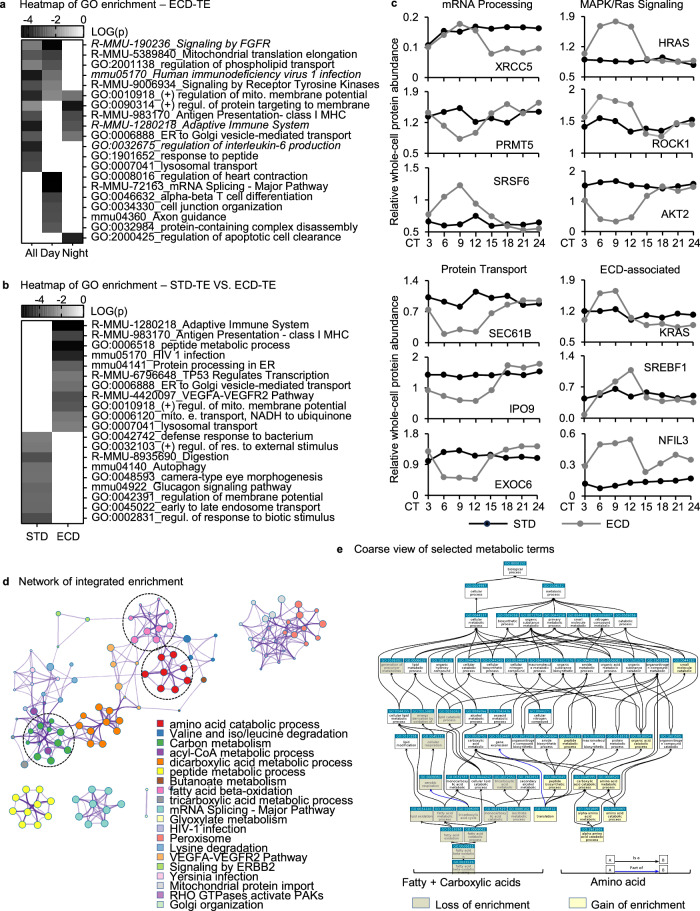
Circadian gene expression is fundamental to the establishment and functions of the circadian clock, a cell-autonomous and evolutionary-conserved timing system. Yet, how it is affected by environmental-circadian disruption (ECD) such as shiftwork and jetlag are ill-defined. Here, we provided a comprehensive and comparative description of male liver circadian gene expression, encompassing transcriptomes, whole-cell proteomes and nuclear proteomes, under normal and after ECD conditions. Under both conditions, post-translation, rather than transcription, is the dominant contributor to circadian functional outputs. After ECD, post-transcriptional and post-translational processes are the major contributors to whole-cell or nuclear circadian proteome, respectively. Furthermore, ECD re-writes the rhythmicity of 64% transcriptome, 98% whole-cell proteome and 95% nuclear proteome. The re-writing, which is associated with changes of circadian regulatory cis-elements, RNA-processing and protein localization, diminishes circadian regulation of fat and carbohydrate metabolism and persists after one week of ECD-recovery.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Environmental circadian disruption re-programs liver circadian gene expression.bioRxiv [Preprint]. 2023 Aug 29:2023.08.28.555175. doi: 10.1101/2023.08.28.555175. bioRxiv. 2023. Update in: Nat Commun. 2024 Jul 1;15(1):5537. doi: 10.1038/s41467-024-49852-3. PMID: 37693605 Free PMC article. Updated. Preprint.
References
-
- Lowrey, P. L. & Takahashi, J. S. Genetics of the mammalian circadian system: Photic entrainment, circadian pacemaker mechanisms, and posttranslational regulation. Annu. Rev. Genet.10.1146/annurev.genet.34.1.533 (2000). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
