

Abnormal HCK/glutamine/autophagy axis promotes endometriosis development by impairing macrophage phagocytosis
- PMID: 38956970
- PMCID: PMC11533058
- DOI: 10.1111/cpr.13702
Abnormal HCK/glutamine/autophagy axis promotes endometriosis development by impairing macrophage phagocytosis
Abstract
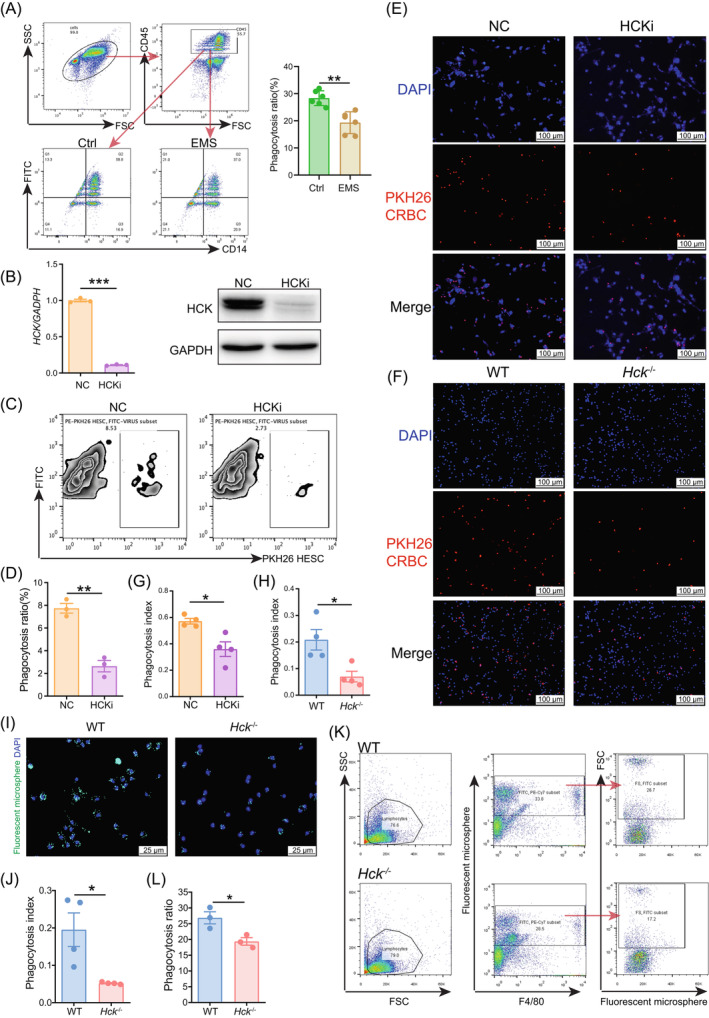
The presence of extensive infiltrated macrophages with impaired phagocytosis is widely recognised as a significant regulator for the development of endometriosis (EMs). Nevertheless, the metabolic characteristics and the fundamental mechanism of impaired macrophage phagocytosis are yet to be clarified. Here, we observe that there is the decreased expression of haematopoietic cellular kinase (HCK) in macrophage of peritoneal fluid from EMs patients, which might be attributed to high oestrogen and hypoxia condition. Of note, HCK deficiency resulted in impaired macrophage phagocytosis, and increased number and weight of ectopic lesions in vitro and in vivo. Mechanistically, this process was mediated via regulation of glutamine metabolism, and further upregulation of macrophage autophagy in a c-FOS/c-JUN dependent manner. Additionally, macrophages of EMs patients displayed insufficient HCK, excessive autophagy and phagocytosis dysfunction. In therapeutic studies, supplementation with glutamine-pre-treated macrophage or Bafilomycin A1 (an autophagy inhibitor)-pre-treated macrophage leads to the induction of macrophage phagocytosis and suppression of EMs development. This observation reveals that the aberrant HCK-glutamine-autophagy axis results in phagocytosis obstacle of macrophage and further increase the development risk of Ems. Additionally, it offers potential therapeutic approaches to prevent EMs, especially patients with insufficient HCK and macrophage phagocytosis dysfunction.
© 2024 The Author(s). Cell Proliferation published by Beijing Institute for Stem Cell and Regenerative Medicine and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures








References
-
- Organization WH . Endometriosis 2023. https://www.who.int/news-room/fact-sheets/detail/endometriosis
-
- Saunders PTK, Horne AW. Endometriosis: etiology, pathobiology, and therapeutic prospects. Cell. 2021;184(11):2807‐2824. - PubMed
-
- Horne AW, Saunders PTK. SnapShot: endometriosis. Cell. 2019;179(7):1677‐1677 e1. - PubMed
-
- Vercellini P, Vigano P, Somigliana E, et al. Endometriosis: pathogenesis and treatment. Nat Rev Endocrinol. 2014;10(5):261‐275. - PubMed
MeSH terms
Substances
Grants and funding
- JIF157602/the Program for Zhuoxue of Fudan University
- IDF157014/002/the Support Project for Original Personalized Research of Fudan University
- 31970798/National Natural Science Foundation of China
- 81771548/National Natural Science Foundation of China
- 82071615/National Natural Science Foundation of China
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
