

The chromatin accessibility and transcriptomic landscape of the aging mice cochlea and the identification of potential functional super-enhancers in age-related hearing loss
- PMID: 38965562
- PMCID: PMC11225416
- DOI: 10.1186/s13148-024-01702-1
The chromatin accessibility and transcriptomic landscape of the aging mice cochlea and the identification of potential functional super-enhancers in age-related hearing loss
Abstract
Background: Presbycusis, also referred to as age-related hearing loss (ARHL), is a condition that results from the cumulative effects of aging on an individual's auditory capabilities. Given the limited understanding of epigenetic mechanisms in ARHL, our research focuses on alterations in chromatin-accessible regions.
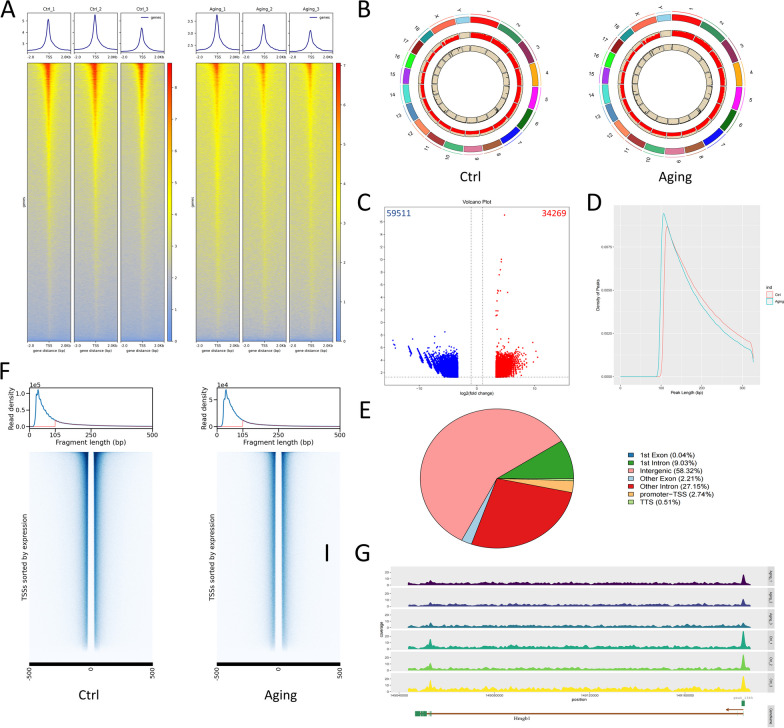
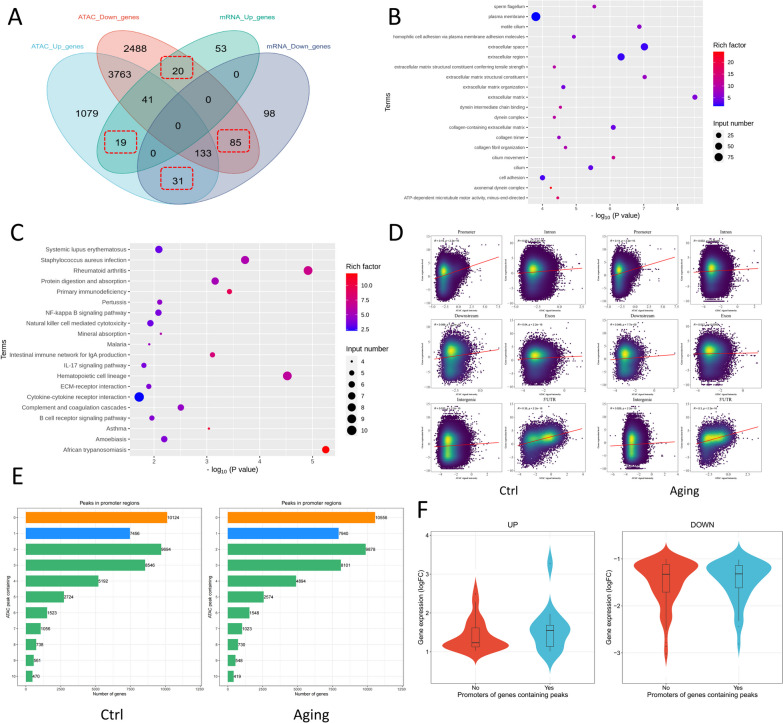
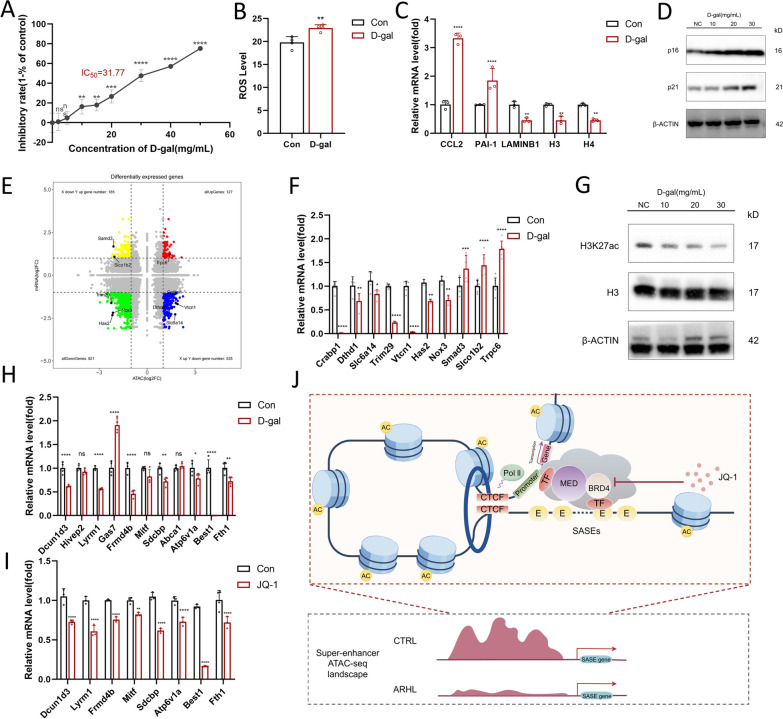
Methods: We employed assay for transposase-accessible chromatin with high-throughput sequencing (ATAC-seq) in conjunction with unique identifier (UID) mRNA-seq between young and aging cochleae, and conducted integrated analysis as well as motif/TF-gene prediction. Additionally, the essential role of super-enhancers (SEs) in the development of ARHL was identified by comparative analysis to previous research. Meanwhile, an ARHL mouse model and an aging mimic hair cell (HC) model were established with a comprehensive identification of senescence phenotypes to access the role of SEs in ARHL progression.
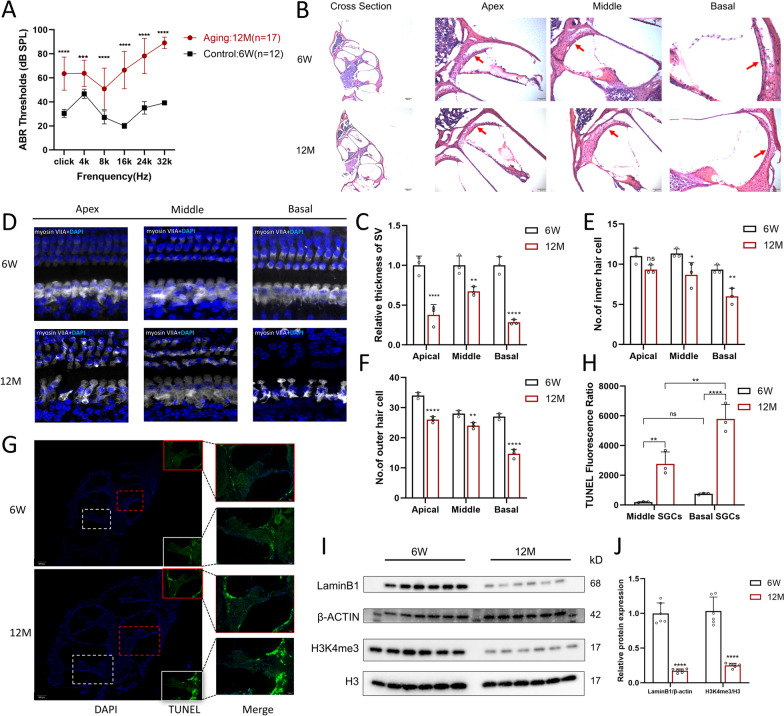
Results: The control cochlear tissue exhibited greater chromatin accessibility than cochlear tissue affected by ARHL. Furthermore, the levels of histone 3 lysine 27 acetylation were significantly depressed in both aging cochlea and aging mimic HEI-OC1 cells, highlighting the essential role of SEs in the development of ARHL. The potential senescence-associated super-enhancers (SASEs) of ARHL were identified, most of which exhibited decreased chromatin accessibility. The majority of genes related to the SASEs showed obvious decreases in mRNA expression level in aging HCs and was noticeably altered following treatment with JQ1 (a commonly used SE inhibitor).
Conclusion: The chromatin accessibility in control cochlear tissue was higher than that in cochlear tissue affected by ARHL. Potential SEs involved in ARHL were identified, which might provide a basis for future therapeutics targeting SASEs related to ARHL.
Keywords: Age-related hearing loss; Chromatin accessibility; Epigenetics; Senescence; Super-enhancer.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
